

Human immunoglobulin repertoire analysis guides design of vaccine priming immunogens targeting HIV V2-apex broadly neutralizing antibody precursors
- PMID: 36179689
- PMCID: PMC9671094
- DOI: 10.1016/j.immuni.2022.09.001
Human immunoglobulin repertoire analysis guides design of vaccine priming immunogens targeting HIV V2-apex broadly neutralizing antibody precursors
Abstract
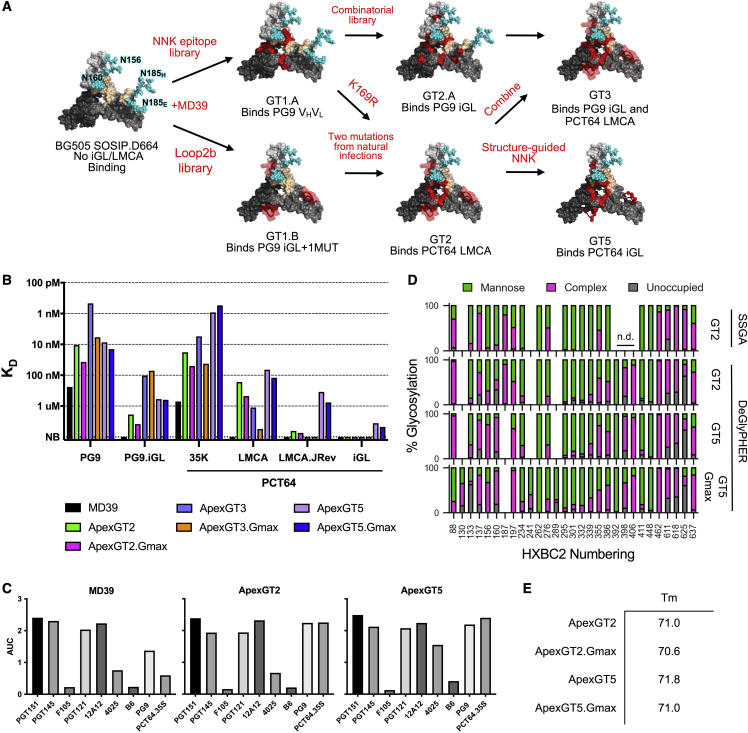
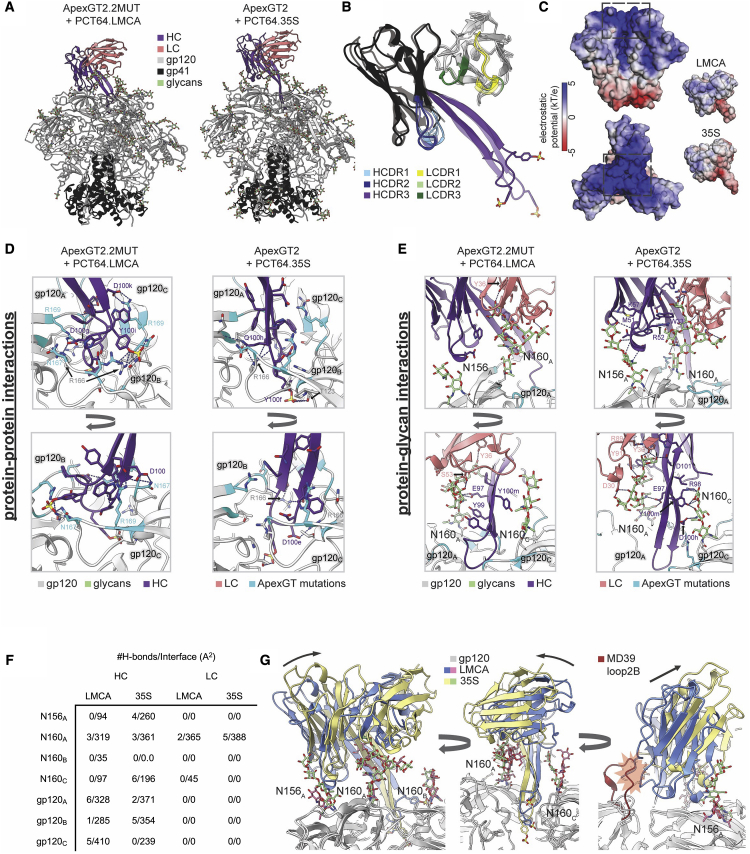
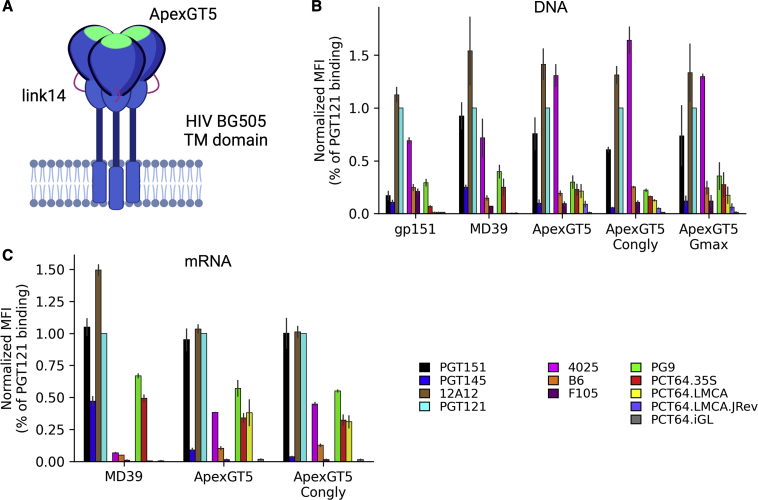
Broadly neutralizing antibodies (bnAbs) to the HIV envelope (Env) V2-apex region are important leads for HIV vaccine design. Most V2-apex bnAbs engage Env with an uncommonly long heavy-chain complementarity-determining region 3 (HCDR3), suggesting that the rarity of bnAb precursors poses a challenge for vaccine priming. We created precursor sequence definitions for V2-apex HCDR3-dependent bnAbs and searched for related precursors in human antibody heavy-chain ultradeep sequencing data from 14 HIV-unexposed donors. We found potential precursors in a majority of donors for only two long-HCDR3 V2-apex bnAbs, PCT64 and PG9, identifying these bnAbs as priority vaccine targets. We then engineered ApexGT Env trimers that bound inferred germlines for PCT64 and PG9 and had higher affinities for bnAbs, determined cryo-EM structures of ApexGT trimers complexed with inferred-germline and bnAb forms of PCT64 and PG9, and developed an mRNA-encoded cell-surface ApexGT trimer. These methods and immunogens have promise to assist HIV vaccine development.
Keywords: AIDS vaccines; HIV antibodies; germline targeting; immunoinformatics; structural vaccinology.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.R.W., K.M.M., J.M.S., and W.R.S. are named inventors on patent applications filed by Scripps and IAVI regarding ApexGT immunogens in this manuscript.
Figures





Similar articles
-
Membrane-bound mRNA immunogens lower the threshold to activate HIV Env V2 apex-directed broadly neutralizing B cell precursors in humanized mice.Immunity. 2022 Nov 8;55(11):2168-2186.e6. doi: 10.1016/j.immuni.2022.09.003. Epub 2022 Sep 29. Immunity. 2022. PMID: 36179690 Free PMC article.
-
Neutralizing Antibody Responses Induced by HIV-1 Envelope Glycoprotein SOSIP Trimers Derived from Elite Neutralizers.J Virol. 2020 Nov 23;94(24):e01214-20. doi: 10.1128/JVI.01214-20. Print 2020 Nov 23. J Virol. 2020. PMID: 32999024 Free PMC article.
-
Antibodies from Rabbits Immunized with HIV-1 Clade B SOSIP Trimers Can Neutralize Multiple Clade B Viruses by Destabilizing the Envelope Glycoprotein.J Virol. 2021 Aug 10;95(17):e0009421. doi: 10.1128/JVI.00094-21. Epub 2021 Aug 10. J Virol. 2021. PMID: 34076487 Free PMC article.
-
Env Exceptionalism: Why Are HIV-1 Env Glycoproteins Atypical Immunogens?Cell Host Microbe. 2020 Apr 8;27(4):507-518. doi: 10.1016/j.chom.2020.03.018. Cell Host Microbe. 2020. PMID: 32272076 Free PMC article. Review.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
Cited by
-
Heavy-chain CDR3-engineered B cells facilitate in vivo evaluation of HIV-1 vaccine candidates.Immunity. 2023 Oct 10;56(10):2408-2424.e6. doi: 10.1016/j.immuni.2023.07.003. Epub 2023 Aug 1. Immunity. 2023. PMID: 37531955 Free PMC article.
-
Molecular basis of SARS-CoV-2 Omicron variant evasion from shared neutralizing antibody response.bioRxiv [Preprint]. 2022 Oct 24:2022.10.24.513517. doi: 10.1101/2022.10.24.513517. bioRxiv. 2022. Update in: Structure. 2023 Jul 6;31(7):801-811.e5. doi: 10.1016/j.str.2023.04.010. PMID: 36324804 Free PMC article. Updated. Preprint.
-
RAIN: a Machine Learning-based identification for HIV-1 bNAbs.Res Sq [Preprint]. 2024 Mar 8:rs.3.rs-4023897. doi: 10.21203/rs.3.rs-4023897/v1. Res Sq. 2024. Update in: Nat Commun. 2024 Jun 24;15(1):5339. doi: 10.1038/s41467-024-49676-1. PMID: 38903123 Free PMC article. Updated. Preprint.
-
RAIN: machine learning-based identification for HIV-1 bNAbs.Nat Commun. 2024 Jun 24;15(1):5339. doi: 10.1038/s41467-024-49676-1. Nat Commun. 2024. PMID: 38914562 Free PMC article.
-
Beyond glycan barriers: non-cognate ligands and protein mimicry approaches to elicit broadly neutralizing antibodies for HIV-1.J Biomed Sci. 2024 Aug 21;31(1):83. doi: 10.1186/s12929-024-01073-y. J Biomed Sci. 2024. PMID: 39169357 Free PMC article. Review.
References
-
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C., et al. Precursor frequency and affinity determine B cell competitive fitness in germinal centers, tested with germline-targeting HIV vaccine immunogens. Immunity. 2018;48:133–146.e6. doi: 10.1016/j.immuni.2017.11.023. - DOI - PMC - PubMed
-
- Agresti A., Coull B.A. Approximate is better than "Exact" for interval estimation of binomial proportions. Am. Statistician. 1998;52:119–126. doi: 10.2307/2685469. - DOI
-
- Alam S.M., Dennison S.M., Aussedat B., Vohra Y., Park P.K., Fernández-Tejada A., Stewart S., Jaeger F.H., Anasti K., Blinn J.H., et al. Recognition of synthetic glycopeptides by HIV-1 broadly neutralizing antibodies and their unmutated ancestors. Proc. Natl. Acad. Sci. USA. 2013;110:18214–18219. doi: 10.1073/pnas.1317855110. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
