

doi: 10.1038/s41594-022-00833-4.
Epub 2022 Sep 12.
Cryo-EM structure of disease-related prion fibrils provides insights into seeding barriers
Affiliations
- PMID: 36097290
- PMCID: PMC9639217
- DOI: 10.1038/s41594-022-00833-4
Item in Clipboard
Cryo-EM structure of disease-related prion fibrils provides insights into seeding barriers
Nat Struct Mol Biol.
2022 Oct.
Abstract
One of the least understood aspects of prion diseases is the structure of infectious prion protein aggregates. Here we report a high-resolution cryo-EM structure of amyloid fibrils formed by human prion protein with the Y145Stop mutation that is associated with a familial prion disease. This structural insight allows us not only to explain previous biochemical findings, but also provides direct support for the conformational adaptability model of prion transmissibility barriers.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
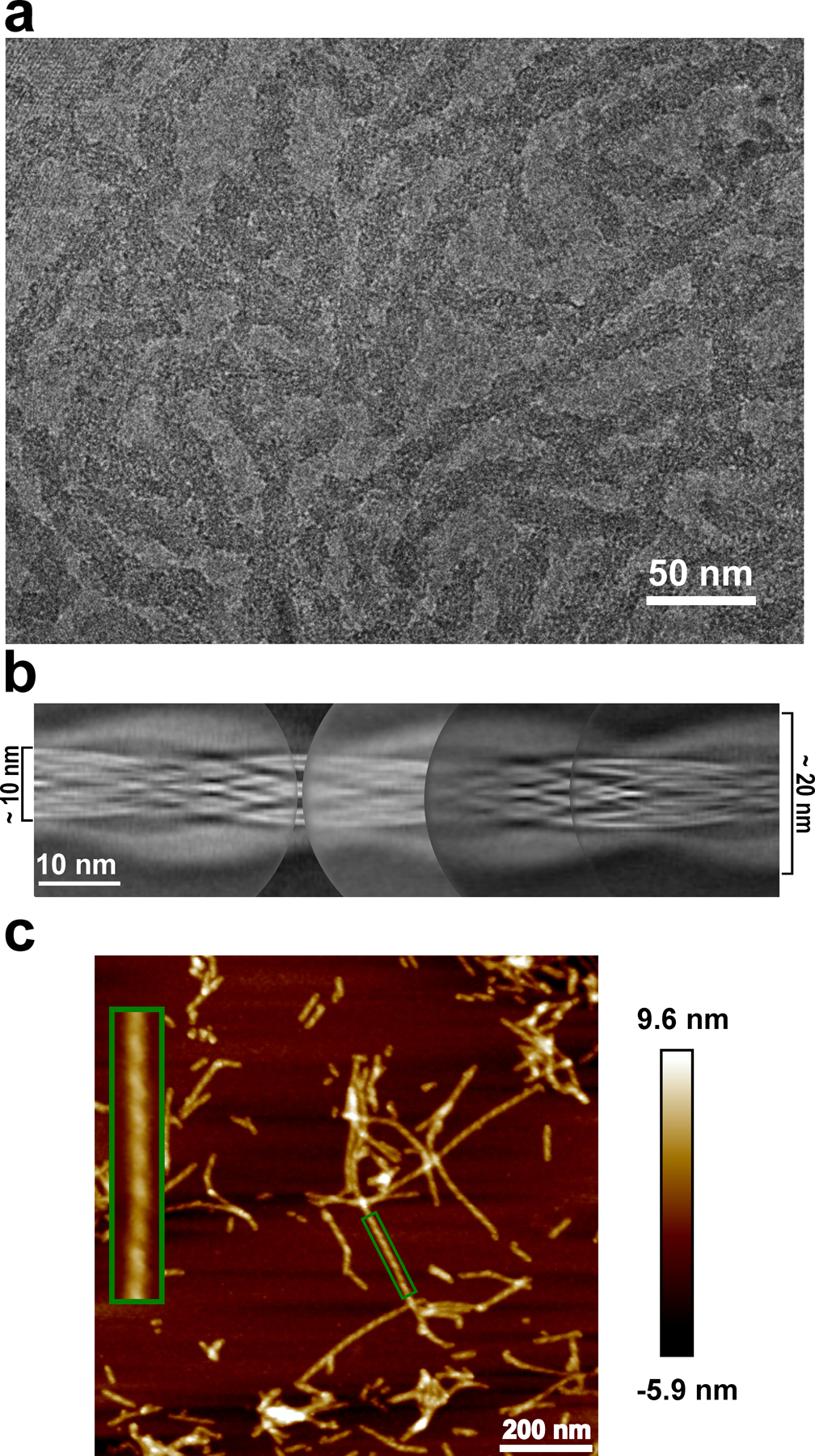
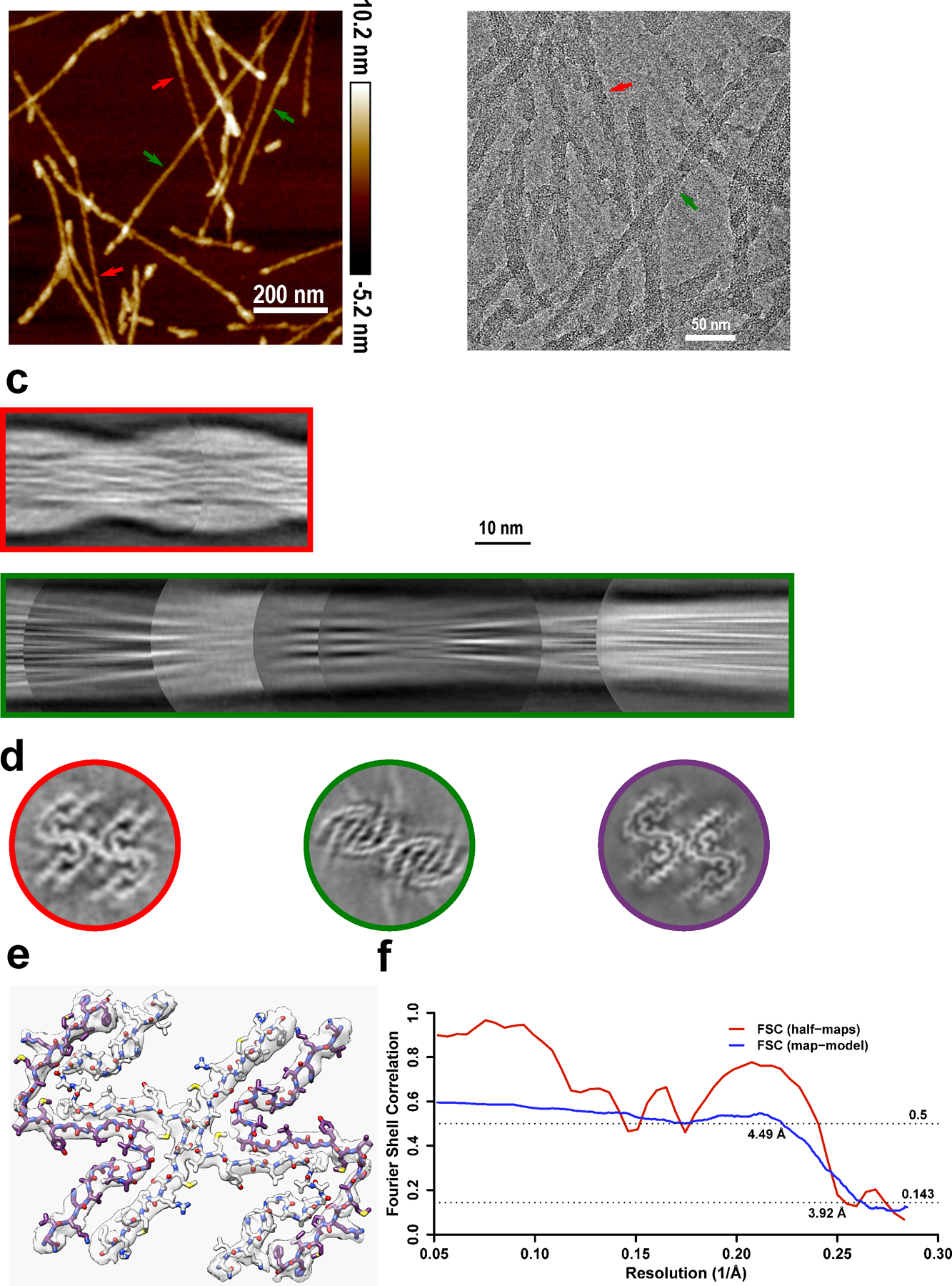
a, Cryo-EM image showing the sole morphology with an apparent twist observed for these fibrils, the same fibril morphology was observed in at least 600 images. b, Manually assembled full pitch of huPrP23–144 fibrils from multiple 2D class averages. The entire fibril has a thickness of ~20 nm and the highly ordered amyloid core has a thickness of ~10 nm. c, AFM image of huPrP23–144 fibrils. A representative fibril in the green box is enlarged to show the left-handed twist, the same fibril morphology was observed in at least 20 images.
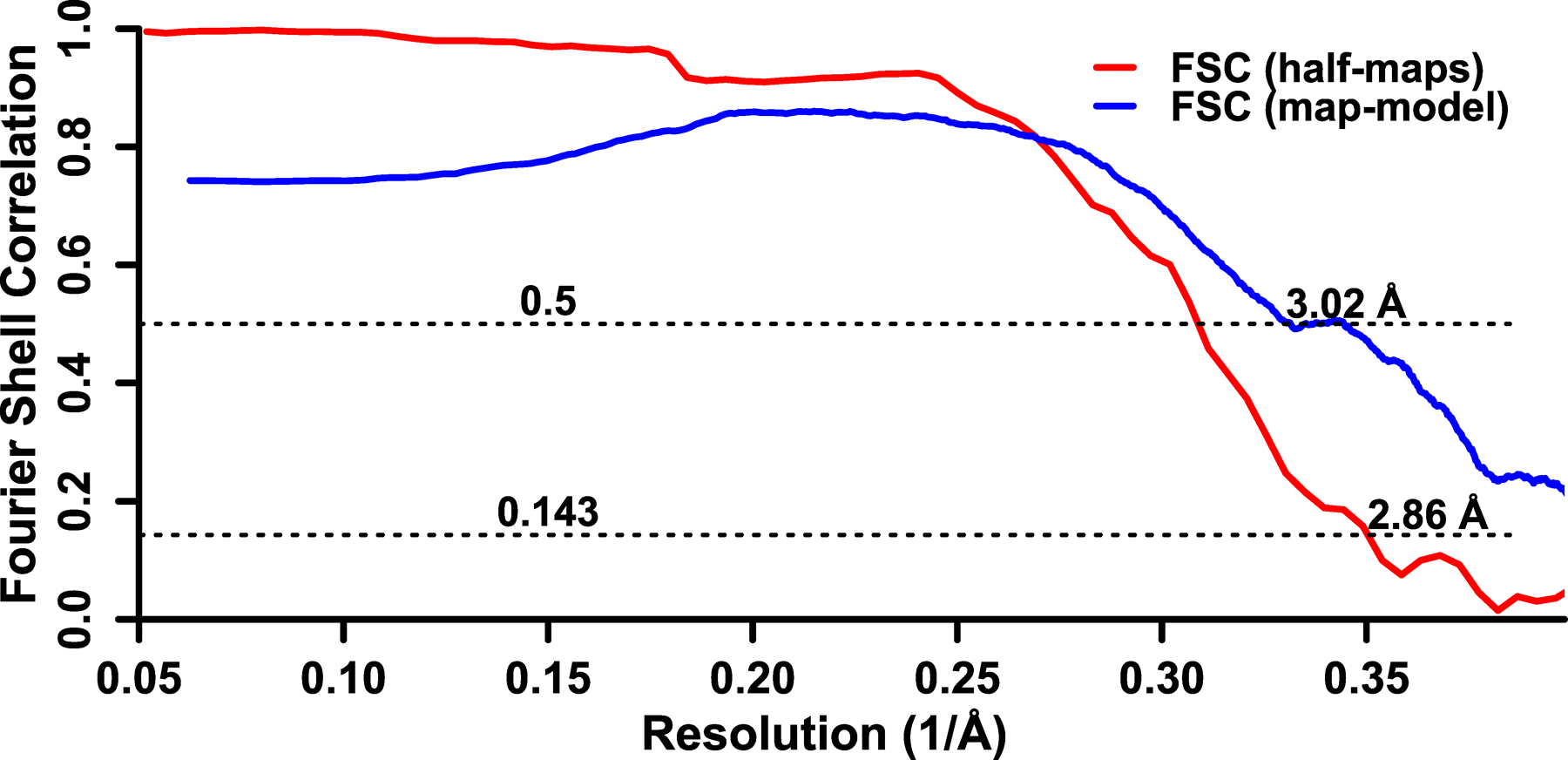
Fourier shell correlation between two independently refined half-maps is shown in red and Fourier shell correlation between the map and the atomic model is shown in blue.
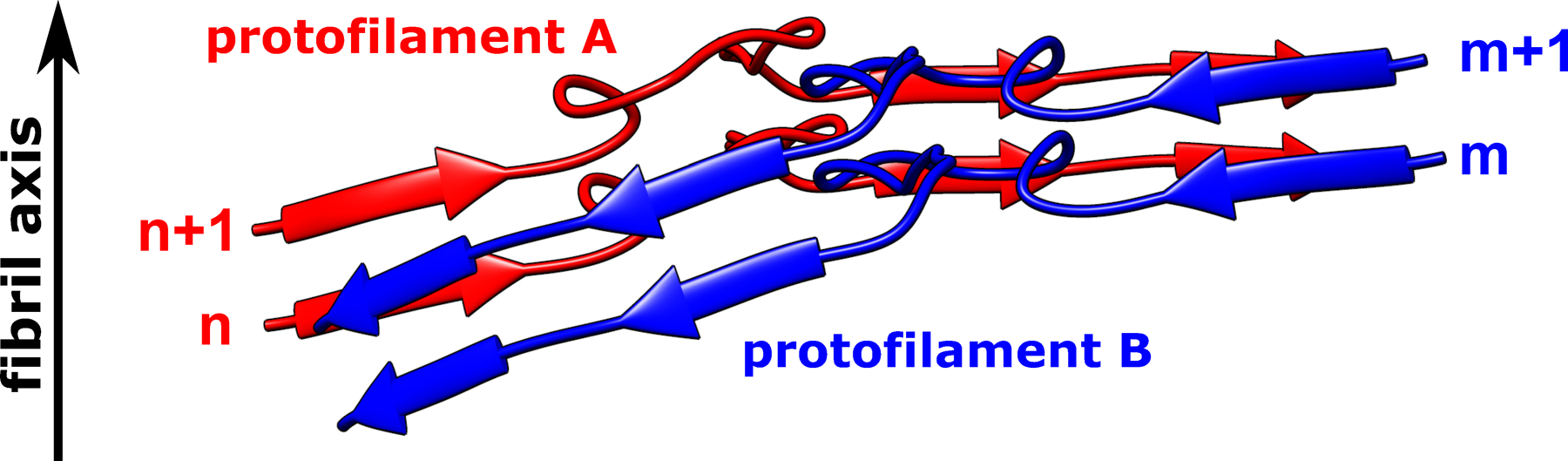
Subunit n in protofilament A interacts with two subunits in protofilament B (subunits m and m+1); subunit m+1 in protofilament B interacts with two subunits in protofilament A (subunits n and n+1). Such a non-planar conformation results in rugged surfaces at fibril ends, with N-terminal residues at the top end and C-terminal residues at the bottom end exposed to water. A similar non-planar assembly is observed for subunits in protofilaments C and D.
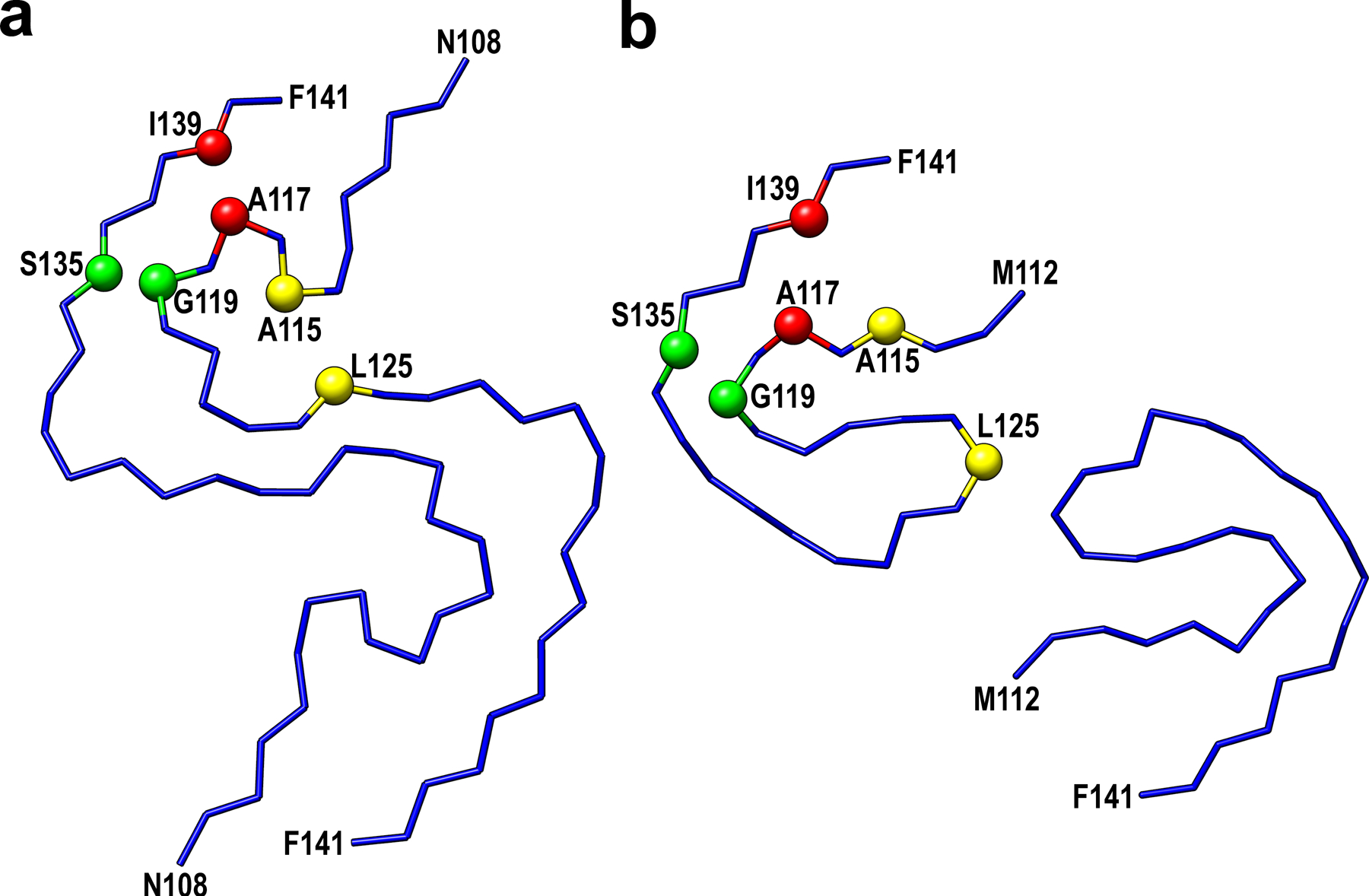
Selected residues involved in close interactions between side-chains in these two models are labeled in red (Ala117-Ile139), yellow (Ala115-Leu125), and green (Gly119-Ser135).
a-b, Two types of polymorphs can be found in AFM images (a) and cryo-EM micrographs (b). Polymorph 1 (red arrows) showed a larger left-handed twist like huPrP23–144 fibrils, and polymorph 2 (green arrows) showed a much smaller right-handed twist. The same fibril morphologies were observed in at least 1,000 cryo-EM images or 20 AFM images for each type of sample. c, Manually assembled half-pitch of both polymorphs from multiple 2D class averages. d, 3D maps of moPrP23–144 and huPrP23–144 fibrils represented by a central slice perpendicular to the fibril axis. Polymorph 1 of moPrP23–144 fibrils (red) showed the same fold as that in huPrP23–144 fibrils (purple). Polymorph 2 of moPrP23–144 fibrils (green) showed a distinctly different fold. e-f, An atomic model built based on the map for polymorph 1 of moPrP23–144 fibrils, with a map resolution of 3.92 Å and model resolution of 4.49 Å. The backbone fold in this model is identical to that in huPrP23–144 fibrils. The structure of polymorph 2 of moPrP23–144 fibrils could not be determined due to poor quality of cryo-EM data for this polymorphic form.
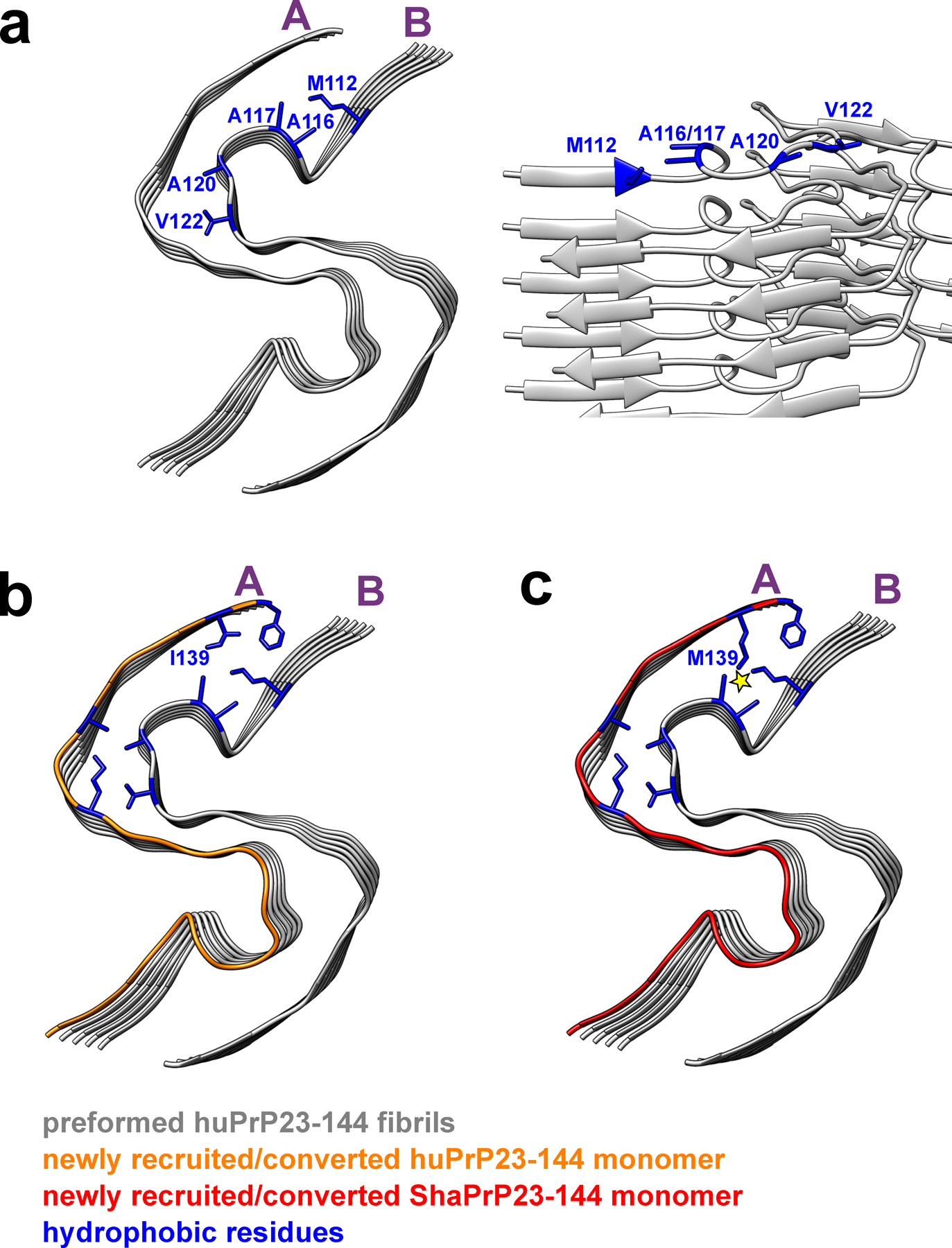
Two representative protofilaments (A and B) are shown only. a, Top and side views of a preformed huPrP23–144 fibril (seed) with solvent-exposed hydrophobic side chains shown in blue. b, Top view of a preformed huPrP23–144 fibril (grey) with a newly recruited and converted subunit of huPrP23–144 (orange). c, Top view of a preformed huPrP23–144 fibril (grey) with a newly recruited subunit of ShaPrP23–144 (red). Adaptation of ShaPrP23–144 to the structure of huPrP23–144 seed would lead to significant intermolecular steric clashes between bulky, elongated side chains of M112 and M139 (as indicated by the yellow star), explaining a cross-seeding barrier.
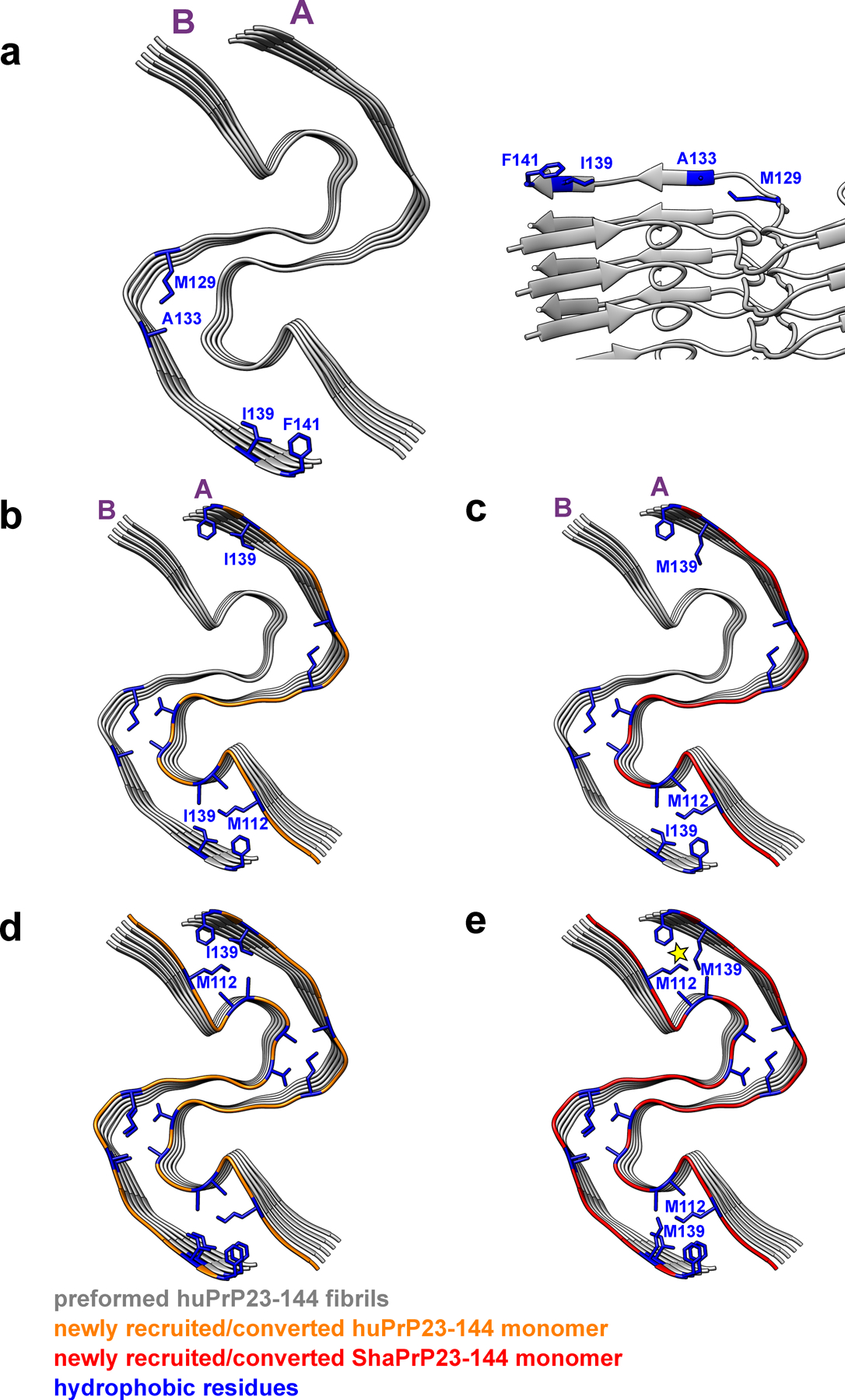
Two representative protofilaments (A and B) are shown for illustrative purposes. a, Bottom and side views of a preformed huPrP23–144 fibril (seed) with solvent-exposed hydrophobic side chains shown in blue. b, Bottom view of a preformed huPrP23–144 fibril (grey) with a newly recruited (to protofilament A) and converted subunit of huPrP23–144 (orange). c, Bottom view of a preformed huPrP23–144 fibril (grey) with a newly recruited (to protofilament A) and converted first subunit of ShaPrP23–144 (red). Due to non-planar structure, C-terminal hydrophobic residues at this end of protofilament B are protruding to water. Thus, recruitment of the first ShaPrP subunit would not result in any steric clashes. d, Bottom view of a preformed huPrP23–144 fibril (grey) with a second newly recruited (to protofilament B) and converted subunit of huPrP23–144 (orange). e, Bottom view of a preformed huPrP23–144 fibril (grey) with a second recruited (to protofilament B) subunit of ShaPrP23–144 (red). Adaptation of this subunit to the structure of the huPrP23–144 seed would lead to intermolecular steric clashes between side chains of M139 and M112 (as indicated by the yellow star), explaining a cross-seeding barrier.
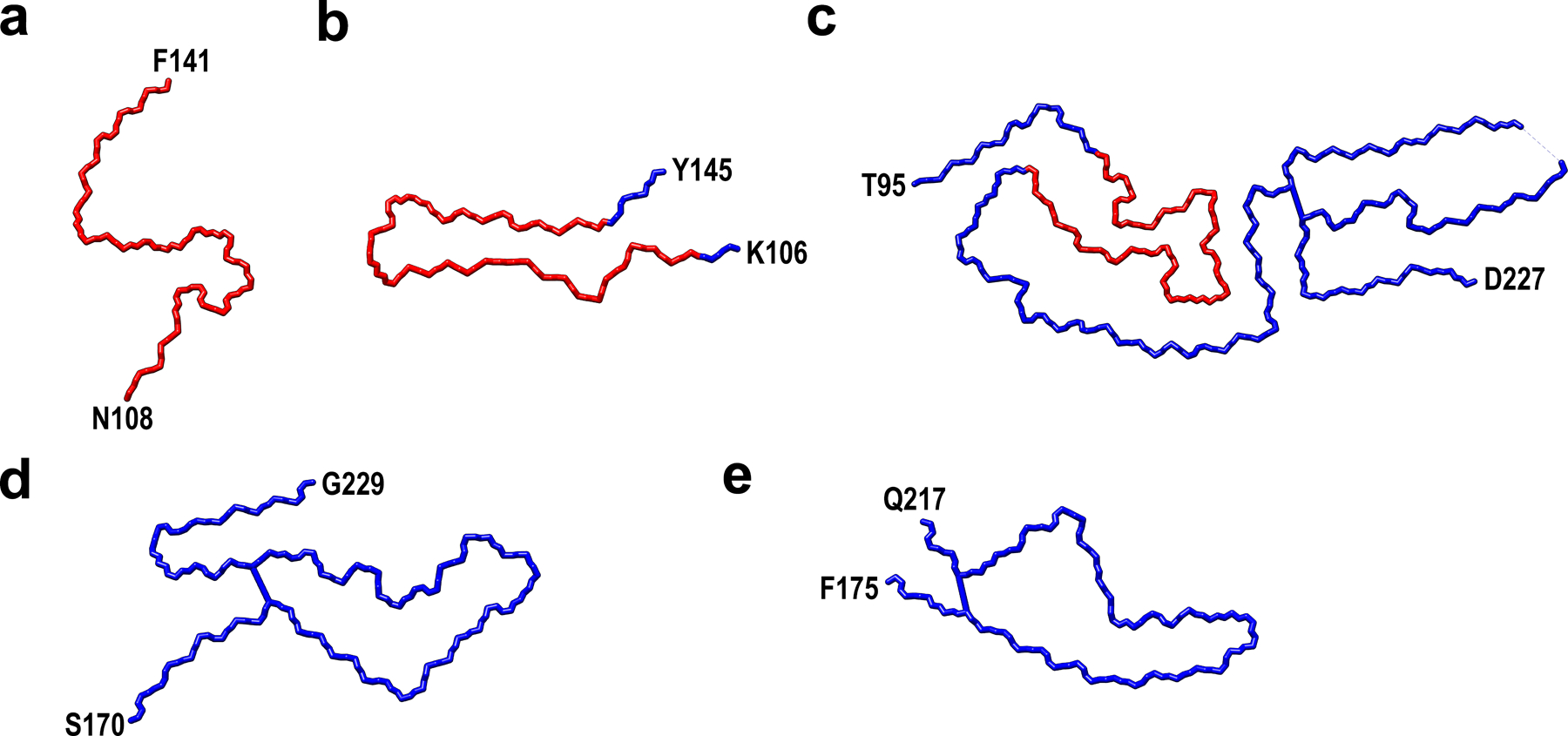
The core of huPrP23–144 fibrils is marked as red.
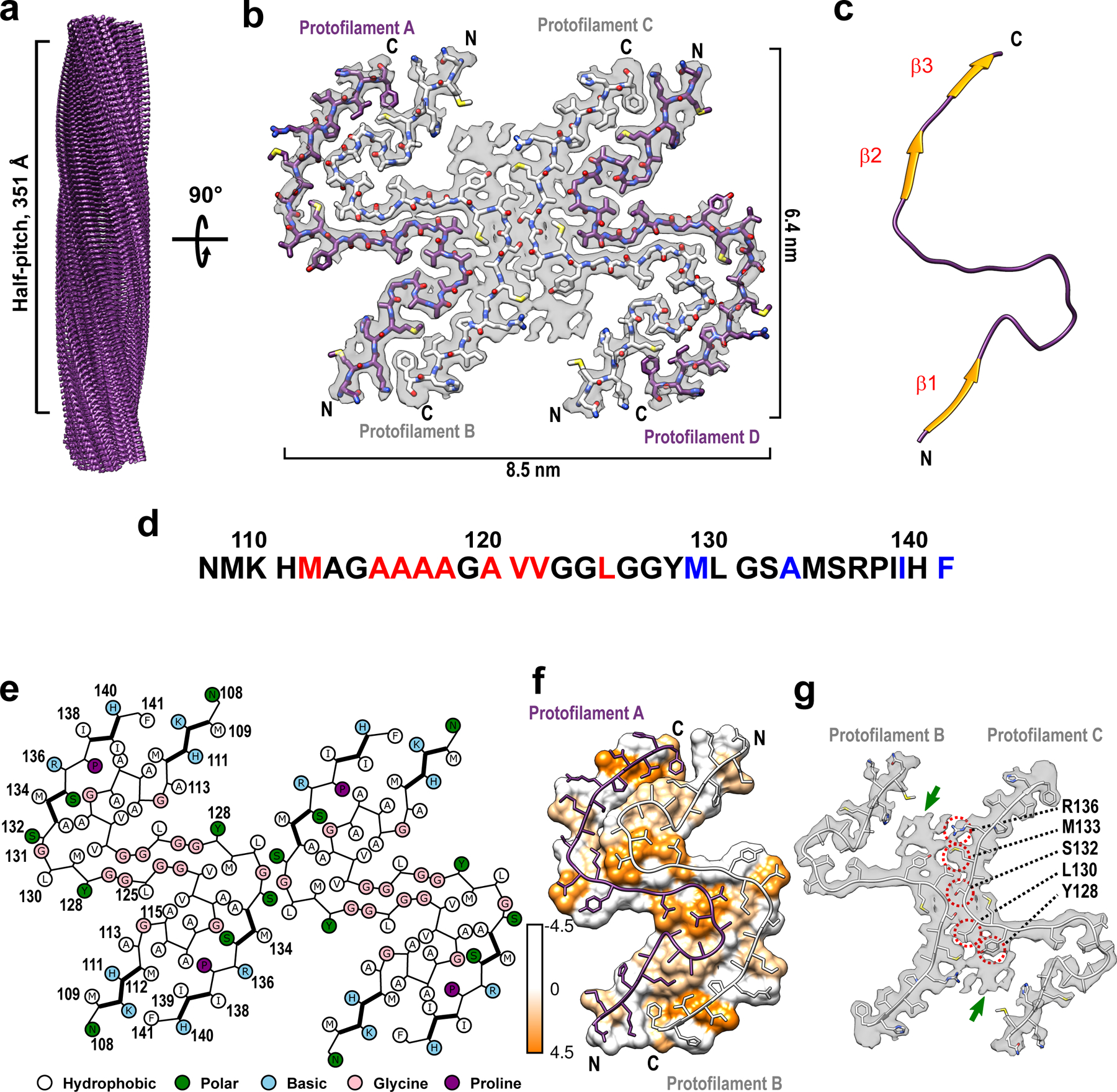
a Cryo-EM density map showing a left-handed twisted helix with a half-pitch of 351 Å and a classical parallel, in-register β-sheet architecture. b The top view of the atomic model superimposed on the cryo-EM density map. The amyloid core contains four identical protofilaments. Two outer protofilaments with greater accessibility to water are depicted in purple; two inner protofilaments are depicted in white. c One representative subunit of the fibril core with β-strands shown in orange. d Amino acid sequence for the highly ordered core of huPrP23–144 fibrils. N- and C-terminal solvent-inaccessible hydrophobic residues are shown in red and blue, respectively. e Schematic representation of one cross-sectional layer of the amyloid core, with β-strands shown as thicker lines. f Hydrophobicity of the cross-section of protofilaments A and B, with hydrophobicity levels colored according to Kyte-Doolittle (top view). g One cross-sectional layer of protofilaments B and C shown as atomic model superimposed on the cryo-EM map (top view). Extra unassigned densities (likely representing side chains of residues outside the core region) are indicated by green arrows; residues within the amyloid core involved in the inter-protofilament interactions are indicated by red circles.
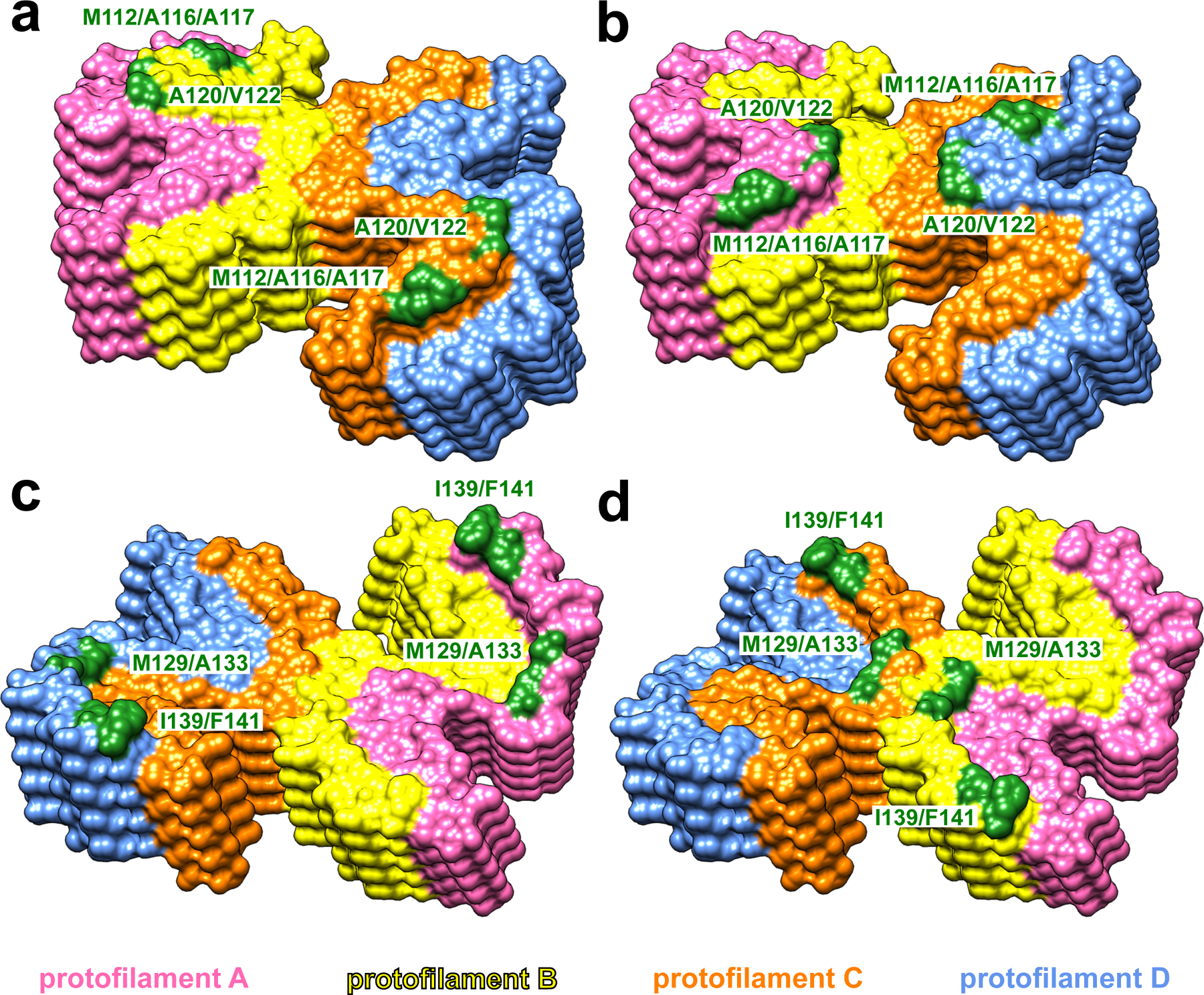
At the top end of the fibril, two clusters of hydrophobic residues (M112/A116/A117 and A120/V122) in protofilaments B and C (yellow and orange, a) or protofilaments A and D (pink and blue, b) are exposed to the solvent. At the bottom end of the fibril, two clusters of hydrophobic residues (M129/A133 and I139/F141) in protofilaments A and D (pink and blue, c) or protofilaments B and C (yellow and orange, d) are exposed to the solvent.
Similar articles
-
PrP P102L and Nearby Lysine Mutations Promote Spontaneous In Vitro Formation of Transmissible Prions.J Virol. 2017 Oct 13;91(21):e01276-17. doi: 10.1128/JVI.01276-17. Print 2017 Nov 1. J Virol. 2017. PMID: 28835493 Free PMC article.
-
The Positively Charged Cluster in the N-terminal Disordered Region may Affect Prion Protein Misfolding: Cryo-EM Structure of Hamster PrP(23-144) Fibrils.J Mol Biol. 2024 Jun 1;436(11):168576. doi: 10.1016/j.jmb.2024.168576. Epub 2024 Apr 18. J Mol Biol. 2024. PMID: 38641239
-
Amyloid fibrils from the N-terminal prion protein fragment are infectious.Proc Natl Acad Sci U S A. 2016 Nov 29;113(48):13851-13856. doi: 10.1073/pnas.1610716113. Epub 2016 Nov 14. Proc Natl Acad Sci U S A. 2016. PMID: 27849581 Free PMC article.
-
The structural line between prion and "prion-like": Insights from prion protein and tau.Curr Opin Neurobiol. 2024 Jun;86:102857. doi: 10.1016/j.conb.2024.102857. Epub 2024 Mar 15. Curr Opin Neurobiol. 2024. PMID: 38489865 Free PMC article. Review.
-
Prion strains viewed through the lens of cryo-EM.Cell Tissue Res. 2023 Apr;392(1):167-178. doi: 10.1007/s00441-022-03676-z. Epub 2022 Aug 27. Cell Tissue Res. 2023. PMID: 36028585 Free PMC article. Review.
Cited by
-
Cryo-EM structures of prion protein filaments from Gerstmann-Sträussler-Scheinker disease.Acta Neuropathol. 2022 Sep;144(3):509-520. doi: 10.1007/s00401-022-02461-0. Epub 2022 Jul 12. Acta Neuropathol. 2022. PMID: 35819518 Free PMC article.
-
Structural biology of ex vivo mammalian prions.J Biol Chem. 2022 Aug;298(8):102181. doi: 10.1016/j.jbc.2022.102181. Epub 2022 Jun 23. J Biol Chem. 2022. PMID: 35752366 Free PMC article. Review.
-
Structural consequences of sequence variation in mammalian prion β2α2 loop segments.Front Neurosci. 2022 Oct 20;16:960322. doi: 10.3389/fnins.2022.960322. eCollection 2022. Front Neurosci. 2022. PMID: 36389229 Free PMC article.
-
Development of a methodology for large-scale production of prions for biological and structural studies.Front Mol Biosci. 2023 Aug 10;10:1184029. doi: 10.3389/fmolb.2023.1184029. eCollection 2023. Front Mol Biosci. 2023. PMID: 37635939 Free PMC article.
-
Pathogenic prion structures at high resolution.PLoS Pathog. 2022 Jun 30;18(6):e1010594. doi: 10.1371/journal.ppat.1010594. eCollection 2022 Jun. PLoS Pathog. 2022. PMID: 35771767 Free PMC article. No abstract available.
References
-
- Kitamoto T, Iizuka R & Tateishi J An amber mutation of prion protein in gerstmann-sträussler syndrome with mutant PrP plaques. Biochemical and Biophysical Research Communications 192, 525–531 (1993). - PubMed
References (to Methods)
-
- Morillas M, Swietnicki W, Gambetti P & Surewicz WK Membrane environment alters the conformational structure of the recombinant human prion protein. Journal of Biological Chemistry 274, 36859–36865 (1999). - PubMed
-
- Mastronarde DN Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology 152, 36–51 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
