

Viral cross-class transmission results in disease of a phytopathogenic fungus
- PMID: 36045287
- PMCID: PMC9428384
- DOI: 10.1038/s41396-022-01310-y
Viral cross-class transmission results in disease of a phytopathogenic fungus
Abstract
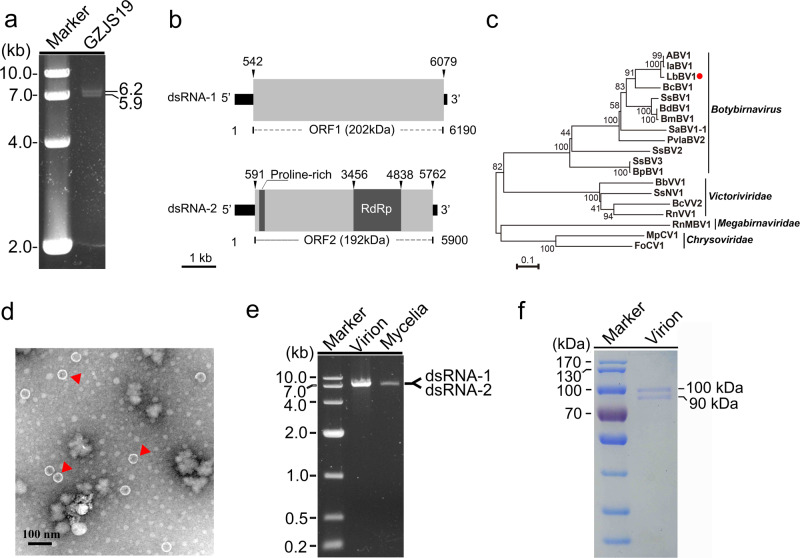
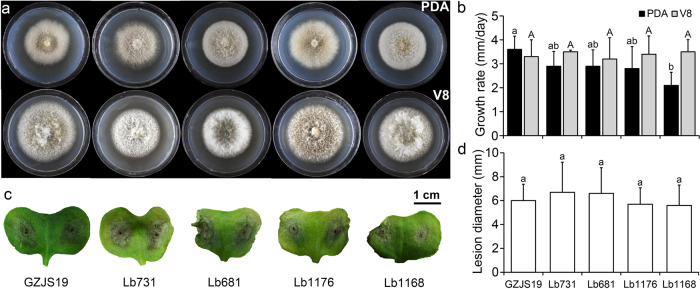
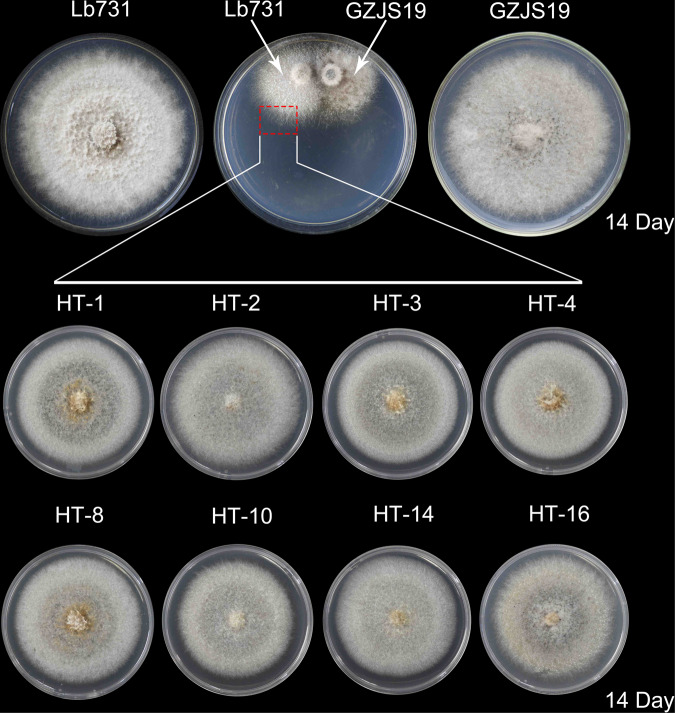
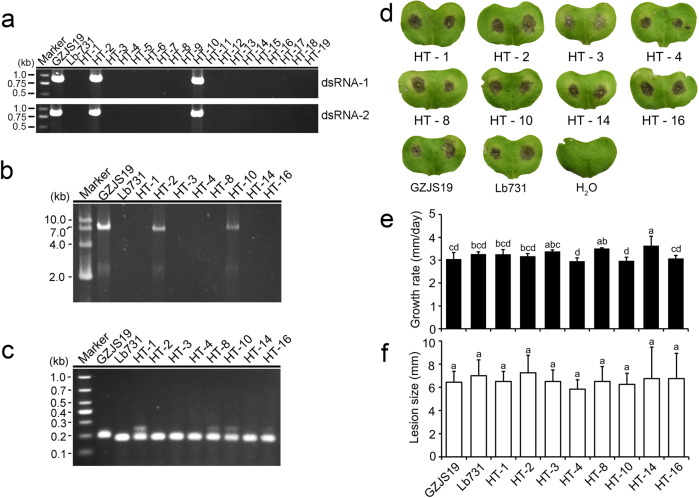
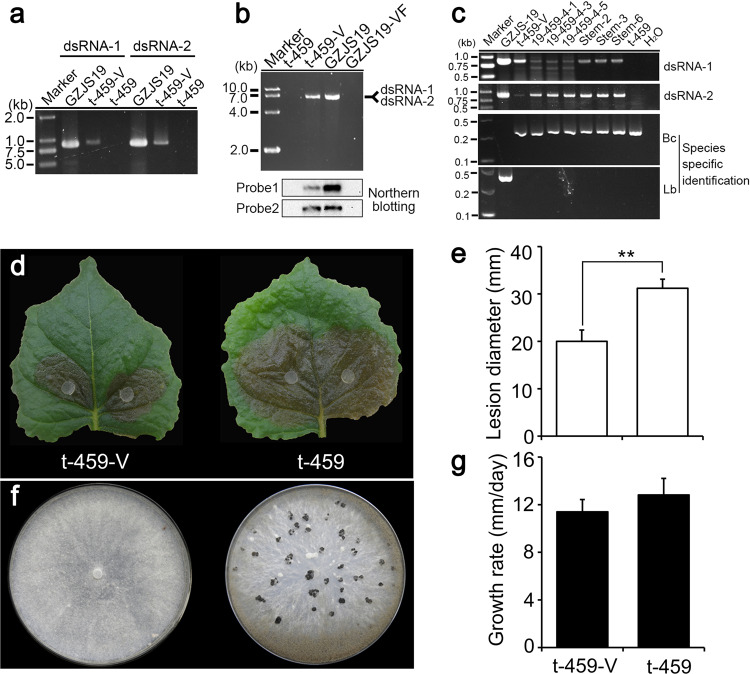
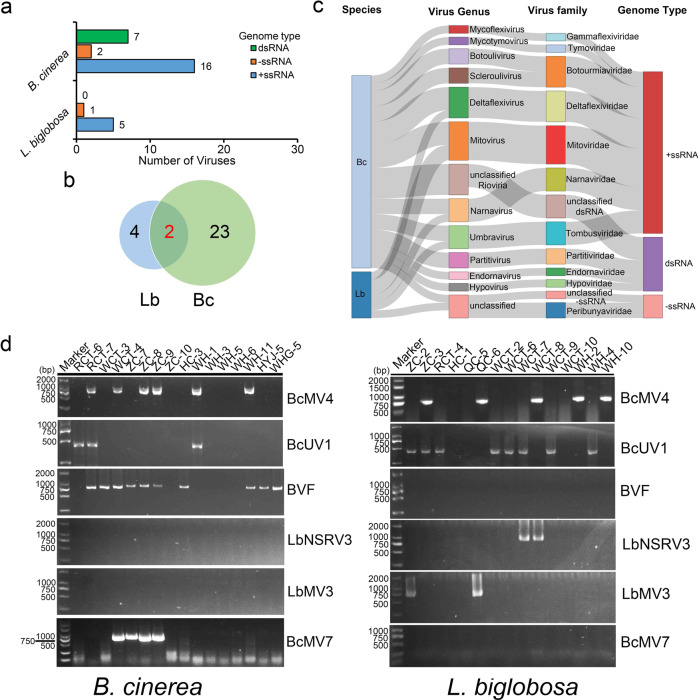
Interspecies transmission of viruses is a well-known phenomenon in animals and plants whether via contacts or vectors. In fungi, interspecies transmission between distantly related fungi is often suspected but rarely experimentally documented and may have practical implications. A newly described double-strand RNA (dsRNA) virus found asymptomatic in the phytopathogenic fungus Leptosphaeria biglobosa of cruciferous crops was successfully transmitted to an evolutionarily distant, broad-host range pathogen Botrytis cinerea. Leptosphaeria biglobosa botybirnavirus 1 (LbBV1) was characterized in L. biglobosa strain GZJS-19. Its infection in L. biglobosa was asymptomatic, as no significant differences in radial mycelial growth and pathogenicity were observed between LbBV1-infected and LbBV1-free strains. However, cross-species transmission of LbBV1 from L. biglobosa to infection in B. cinerea resulted in the hypovirulence of the recipient B. cinerea strain t-459-V. The cross-species transmission was succeeded only by inoculation of mixed spores of L. biglobosa and B. cinerea on PDA or on stems of oilseed rape with the efficiency of 4.6% and 18.8%, respectively. To investigate viral cross-species transmission between L. biglobosa and B. cinerea in nature, RNA sequencing was carried out on L. biglobosa and B. cinerea isolates obtained from Brassica samples co-infected by these two pathogens and showed that at least two mycoviruses were detected in both fungal groups. These results indicate that cross-species transmission of mycoviruses may occur frequently in nature and result in the phenotypical changes of newly invaded phytopathogenic fungi. This study also provides new insights for using asymptomatic mycoviruses as biocontrol agent.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Characterization of a novel RNA virus from the phytopathogenic fungus Leptosphaeria biglobosa related to members of the genus Mitovirus.Arch Virol. 2019 Mar;164(3):913-916. doi: 10.1007/s00705-018-04143-3. Epub 2019 Jan 17. Arch Virol. 2019. PMID: 30656466
-
Identification, Molecular Characterization, and Biology of a Novel Quadrivirus Infecting the Phytopathogenic Fungus Leptosphaeria biglobosa.Viruses. 2018 Dec 25;11(1):9. doi: 10.3390/v11010009. Viruses. 2018. PMID: 30585188 Free PMC article.
-
Co-infection of a hypovirulent isolate of Sclerotinia sclerotiorum with a new botybirnavirus and a strain of a mitovirus.Virol J. 2016 Jun 6;13:92. doi: 10.1186/s12985-016-0550-2. Virol J. 2016. PMID: 27267756 Free PMC article.
-
Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses.Viruses. 2023 May 19;15(5):1202. doi: 10.3390/v15051202. Viruses. 2023. PMID: 37243288 Free PMC article. Review.
-
Viruses of botrytis.Adv Virus Res. 2013;86:249-72. doi: 10.1016/B978-0-12-394315-6.00009-X. Adv Virus Res. 2013. PMID: 23498909 Review.
Cited by
-
Off to a new host.Nat Rev Microbiol. 2022 Nov;20(11):638. doi: 10.1038/s41579-022-00803-z. Nat Rev Microbiol. 2022. PMID: 36127515 No abstract available.
-
Virome release of an invasive exotic plant species in southern France.Virus Evol. 2024 Mar 9;10(1):veae025. doi: 10.1093/ve/veae025. eCollection 2024. Virus Evol. 2024. PMID: 38566975 Free PMC article.
-
Hypovirulence-associated mycovirus epidemics cause pathogenicity degeneration of Beauveria bassiana in the field.Virol J. 2023 Nov 3;20(1):255. doi: 10.1186/s12985-023-02217-6. Virol J. 2023. PMID: 37924080 Free PMC article.
-
Plants interfere with non-self recognition of a phytopathogenic fungus via proline accumulation to facilitate mycovirus transmission.Nat Commun. 2024 Jun 4;15(1):4748. doi: 10.1038/s41467-024-49110-6. Nat Commun. 2024. PMID: 38834585 Free PMC article.
-
Greetings from virologists to mycologists: A review outlining viruses that live in fungi.Mycoscience. 2024 Jan 31;65(1):1-11. doi: 10.47371/mycosci.2023.11.004. eCollection 2024. Mycoscience. 2024. PMID: 39239117 Free PMC article.
References
-
- May RM, Anderson RM. Epidemiology and genetics in the coevolution of parasites and hosts. Proc R Soc B. 1983;219:281–313. - PubMed
-
- Ewald PW. Transmission modes and evolution of the parasitism-mutualism continuum. Ann NY Acad Sci. 1987;503:295–306. - PubMed
-
- Brown MJF, Loosli R, Schmid-Hempel P. Condition-dependent expression of virulence in a trypanosome infecting bumblebees. Oikos. 2000;91:421–7.
