

CD8 coreceptor engagement of MR1 enhances antigen responsiveness by human MAIT and other MR1-reactive T cells
- PMID: 36018322
- PMCID: PMC9424912
- DOI: 10.1084/jem.20210828
CD8 coreceptor engagement of MR1 enhances antigen responsiveness by human MAIT and other MR1-reactive T cells
Abstract
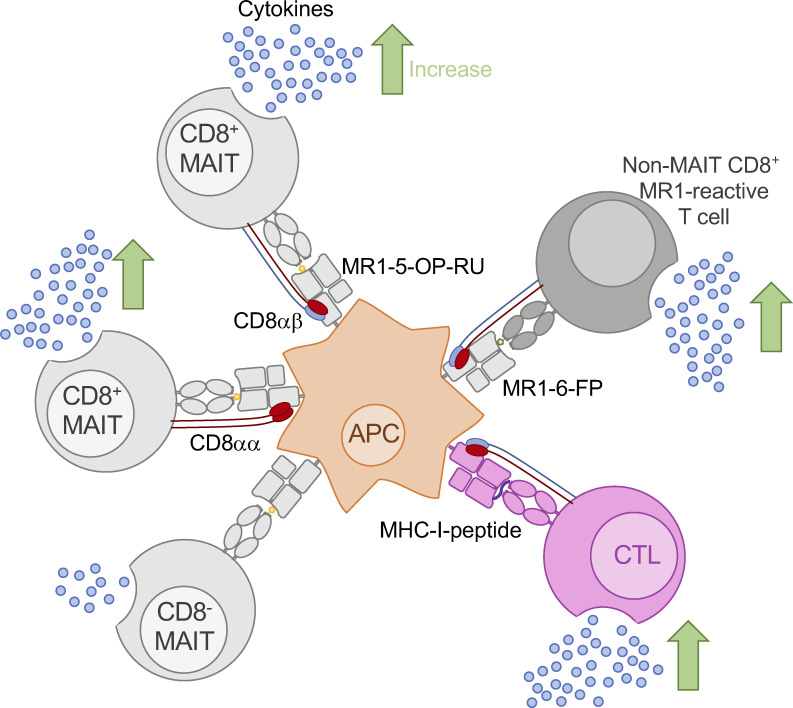
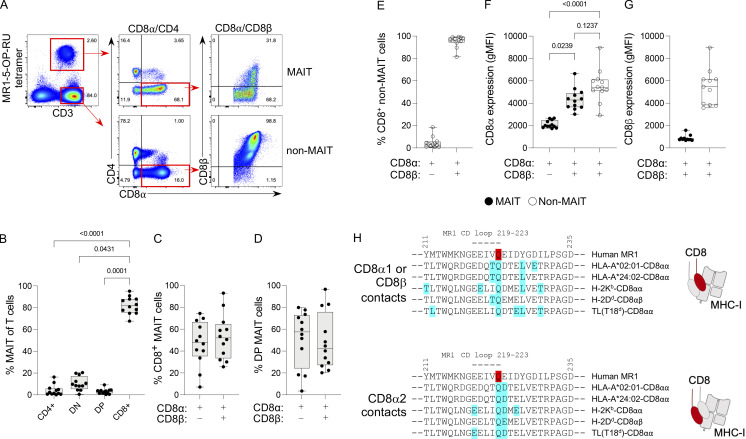
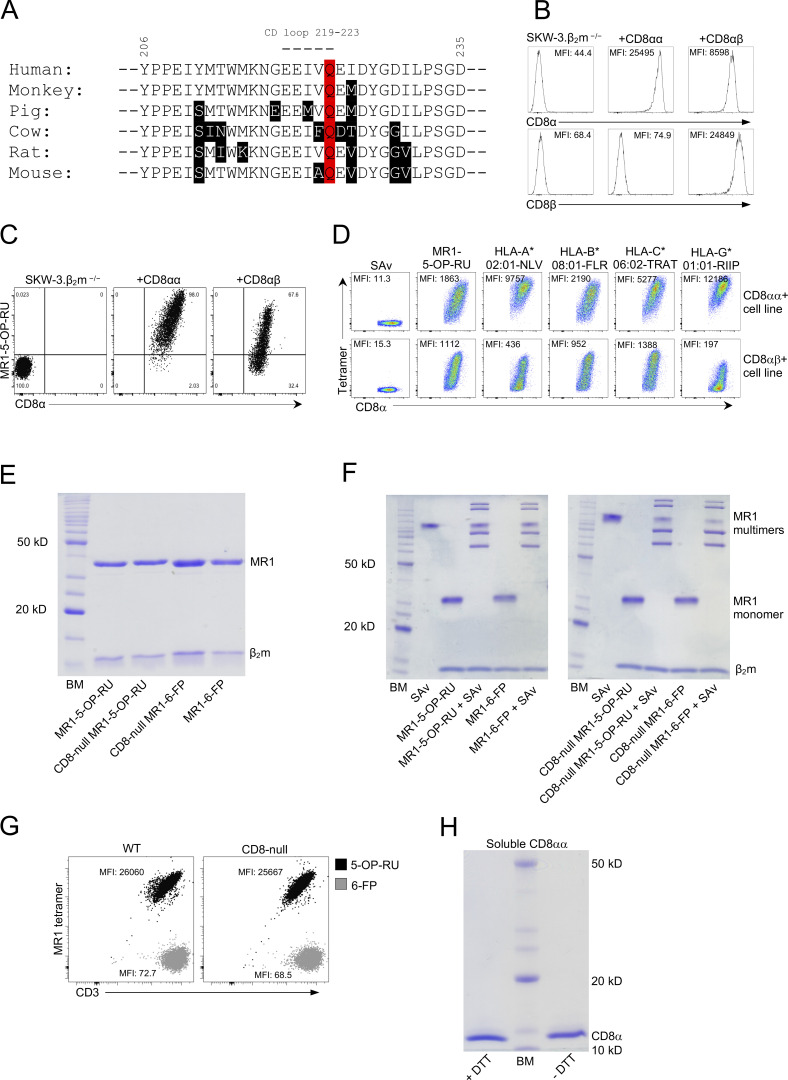
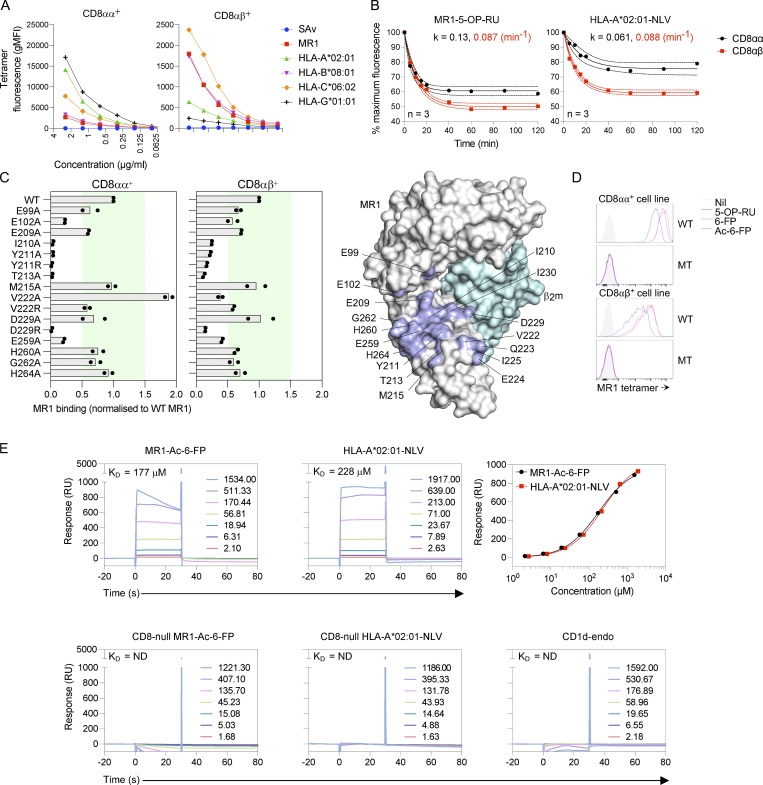
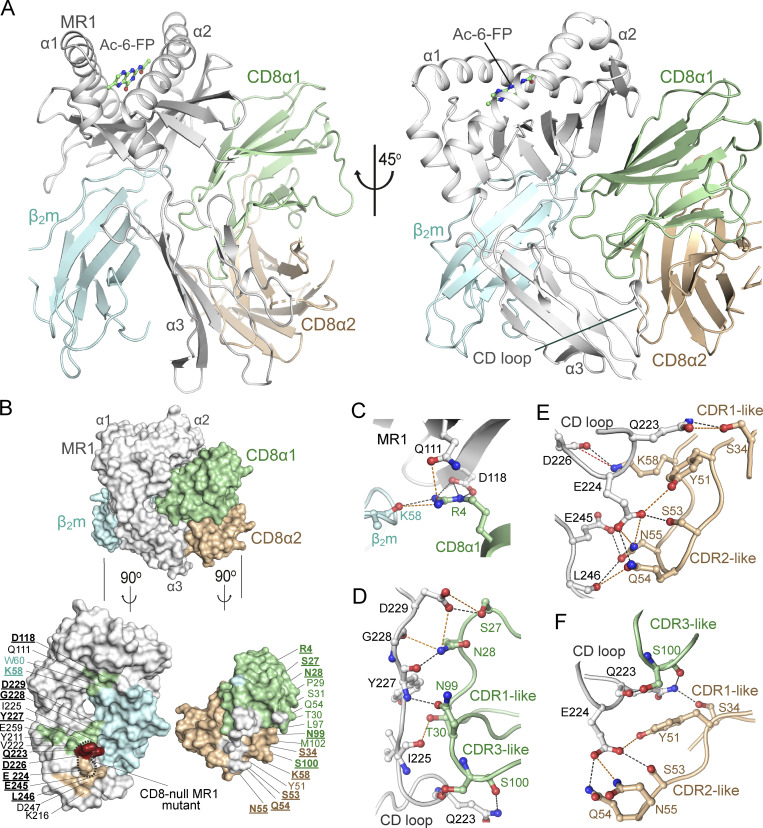
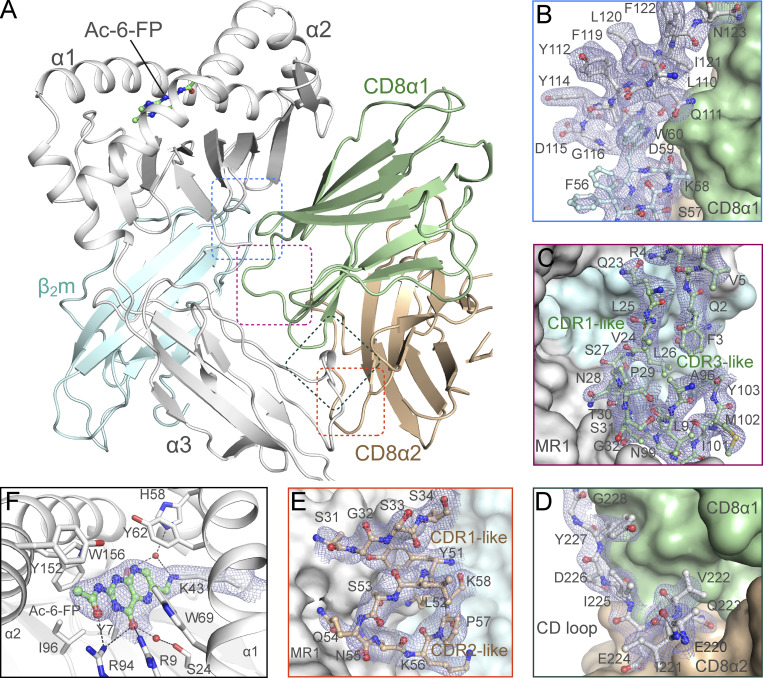
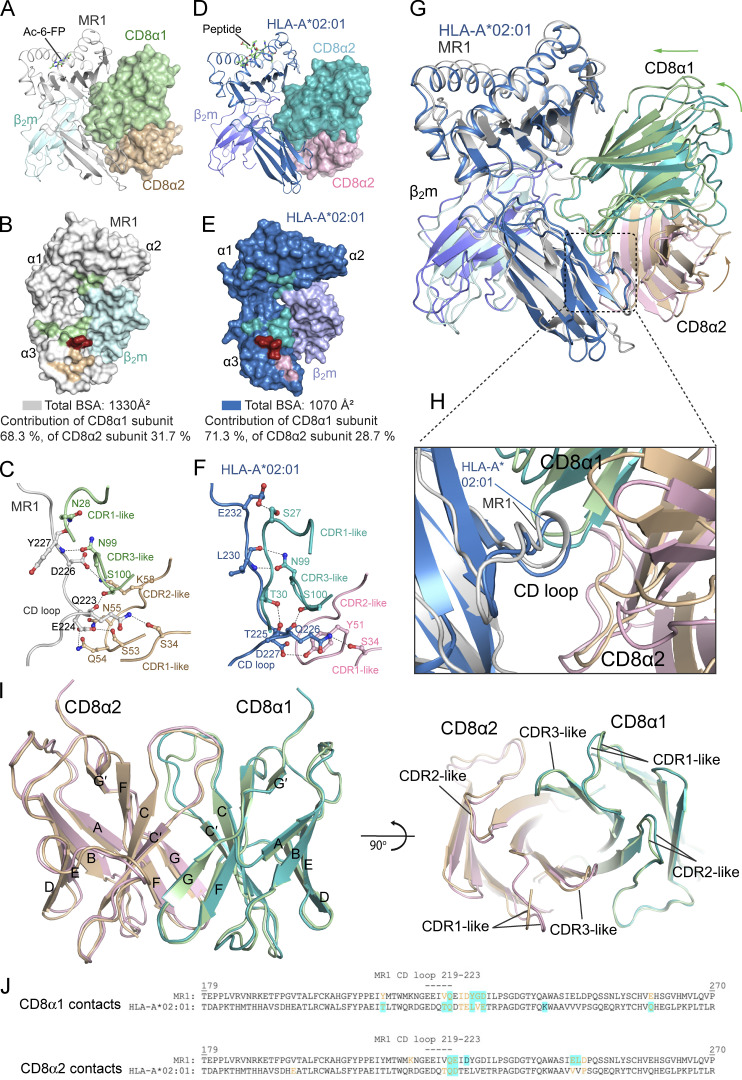
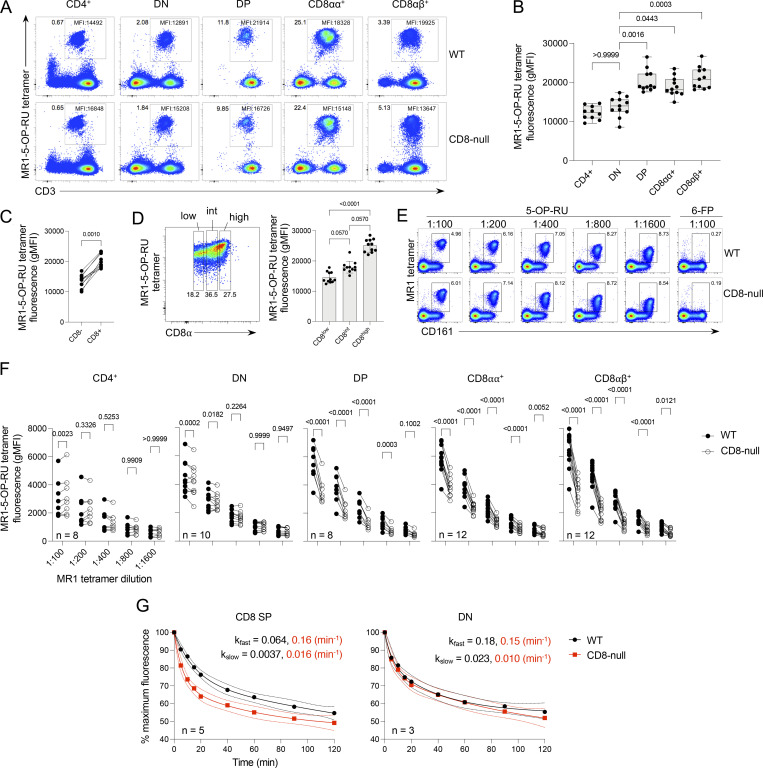
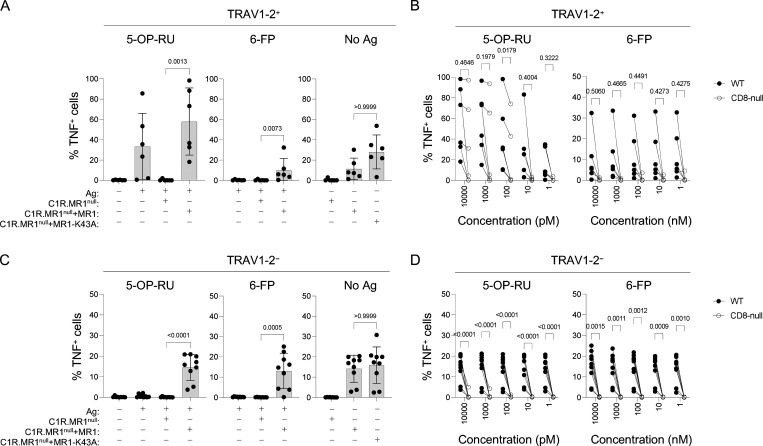
Mucosal-associated invariant T (MAIT) cells detect microbial infection via recognition of riboflavin-based antigens presented by the major histocompatibility complex class I (MHC-I)-related protein 1 (MR1). Most MAIT cells in human peripheral blood express CD8αα or CD8αβ coreceptors, and the binding site for CD8 on MHC-I molecules is relatively conserved in MR1. Yet, there is no direct evidence of CD8 interacting with MR1 or the functional consequences thereof. Similarly, the role of CD8αα in lymphocyte function remains ill-defined. Here, using newly developed MR1 tetramers, mutated at the CD8 binding site, and by determining the crystal structure of MR1-CD8αα, we show that CD8 engaged MR1, analogous to how it engages MHC-I molecules. CD8αα and CD8αβ enhanced MR1 binding and cytokine production by MAIT cells. Moreover, the CD8-MR1 interaction was critical for the recognition of folate-derived antigens by other MR1-reactive T cells. Together, our findings suggest that both CD8αα and CD8αβ act as functional coreceptors for MAIT and other MR1-reactive T cells.
© 2022 The University of Melbourne.
Conflict of interest statement
Disclosures: J.Y.W. Mak, L. Liu, A.J. Corbett, D.P. Fairlie, Z. Chen, J. Rossjohn, J. McCluskey, and S.B.G. Eckle reported a patent (WO/2015/149130) with royalties paid by Immudex and licensed to the NIH Tetramer Core Facility. L. Liu, A.J. Corbett, D.P. Fairlie, J. Rossjohn, J. McCluskey, and S.B.G. Eckle reported a patent (WO/2014/005194) with royalties paid by Immudex and licensed to the NIH Tetramer Core Facility. D.I. Godfrey reported a patent to PROVAU2022900574 pending. No other disclosures were reported.
Figures
Similar articles
-
Antigen-loaded MR1 tetramers define T cell receptor heterogeneity in mucosal-associated invariant T cells.J Exp Med. 2013 Oct 21;210(11):2305-20. doi: 10.1084/jem.20130958. Epub 2013 Oct 7. J Exp Med. 2013. PMID: 24101382 Free PMC article.
-
Functional Heterogeneity and Antimycobacterial Effects of Mouse Mucosal-Associated Invariant T Cells Specific for Riboflavin Metabolites.J Immunol. 2015 Jul 15;195(2):587-601. doi: 10.4049/jimmunol.1402545. Epub 2015 Jun 10. J Immunol. 2015. PMID: 26063000 Free PMC article.
-
MHC class I-related molecule, MR1, and mucosal-associated invariant T cells.Immunol Rev. 2016 Jul;272(1):120-38. doi: 10.1111/imr.12423. Immunol Rev. 2016. PMID: 27319347 Review.
-
A molecular basis underpinning the T cell receptor heterogeneity of mucosal-associated invariant T cells.J Exp Med. 2014 Jul 28;211(8):1585-600. doi: 10.1084/jem.20140484. Epub 2014 Jul 21. J Exp Med. 2014. PMID: 25049336 Free PMC article.
-
Mucosal-associated invariant T cell receptor recognition of small molecules presented by MR1.Immunol Cell Biol. 2018 Jul;96(6):588-597. doi: 10.1111/imcb.12017. Epub 2018 Feb 27. Immunol Cell Biol. 2018. PMID: 29393543 Review.
Cited by
-
Structure, function, and immunomodulation of the CD8 co-receptor.Front Immunol. 2024 Aug 26;15:1412513. doi: 10.3389/fimmu.2024.1412513. eCollection 2024. Front Immunol. 2024. PMID: 39253084 Free PMC article. Review.
-
Molecular insights into metabolite antigen recognition by mucosal-associated invariant T cells.Curr Opin Immunol. 2023 Aug;83:102351. doi: 10.1016/j.coi.2023.102351. Epub 2023 Jun 3. Curr Opin Immunol. 2023. PMID: 37276819 Free PMC article. Review.
-
MAITabolism2 - the emerging understanding of MAIT cell metabolism and their role in metabolic disease.Front Immunol. 2023 Jan 19;13:1108071. doi: 10.3389/fimmu.2022.1108071. eCollection 2022. Front Immunol. 2023. PMID: 36741413 Free PMC article. Review.
-
Antibody-mediated depletion of select leukocyte subsets in blood and tissue of nonhuman primates.Front Immunol. 2024 Mar 11;15:1359679. doi: 10.3389/fimmu.2024.1359679. eCollection 2024. Front Immunol. 2024. PMID: 38529287 Free PMC article.
-
MR1 antigen presentation to MAIT cells and other MR1-restricted T cells.Nat Rev Immunol. 2024 Mar;24(3):178-192. doi: 10.1038/s41577-023-00934-1. Epub 2023 Sep 29. Nat Rev Immunol. 2024. PMID: 37773272 Free PMC article. Review.
References
-
- Adams, P.D., Afonine P.V., Bunkóczi G., Chen V.B., Davis I.W., Echols N., Headd J.J., Hung L.-W., Kapral G.J., Grosse-Kunstleve R.W., et al. . 2010. PHENIX: A comprehensive python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66:213–221. 10.1107/S0907444909052925 - DOI - PMC - PubMed
-
- Allan, D.S., Colonna M., Lanier L.L., Churakova T.D., Abrams J.S., Ellis S.A., Mcmichael A.J., and Braud V.M.. 1999. Tetrameric complexes of human histocompatibility leukocyte antigen (HLA)-G bind to peripheral blood myelomonocytic cells. J. Exp. Med. 189:1149–1156. 10.1084/jem.189.7.1149 - DOI - PMC - PubMed
-
- Aragão, D., Aishima J., Cherukuvada H., Clarken R., Clift M., Cowieson N.P., Ericsson D.J., Gee C.L., Macedo S., Mudie N., et al. . 2018. MX2: A high-flux undulator microfocus beamline serving both the chemical and macromolecular crystallography communities at the Australian Synchrotron. J. Synchrotron Radiat. 25:885–891. 10.1107/S1600577518003120 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
