

Single-cell analyses reveal early thymic progenitors and pre-B cells in zebrafish
- PMID: 35938989
- PMCID: PMC9365674
- DOI: 10.1084/jem.20220038
Single-cell analyses reveal early thymic progenitors and pre-B cells in zebrafish
Abstract
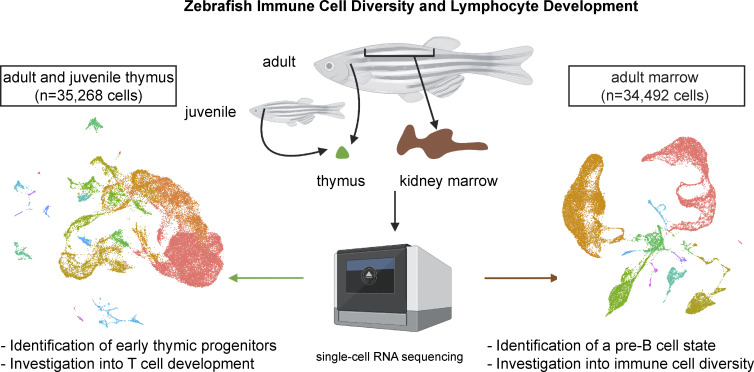
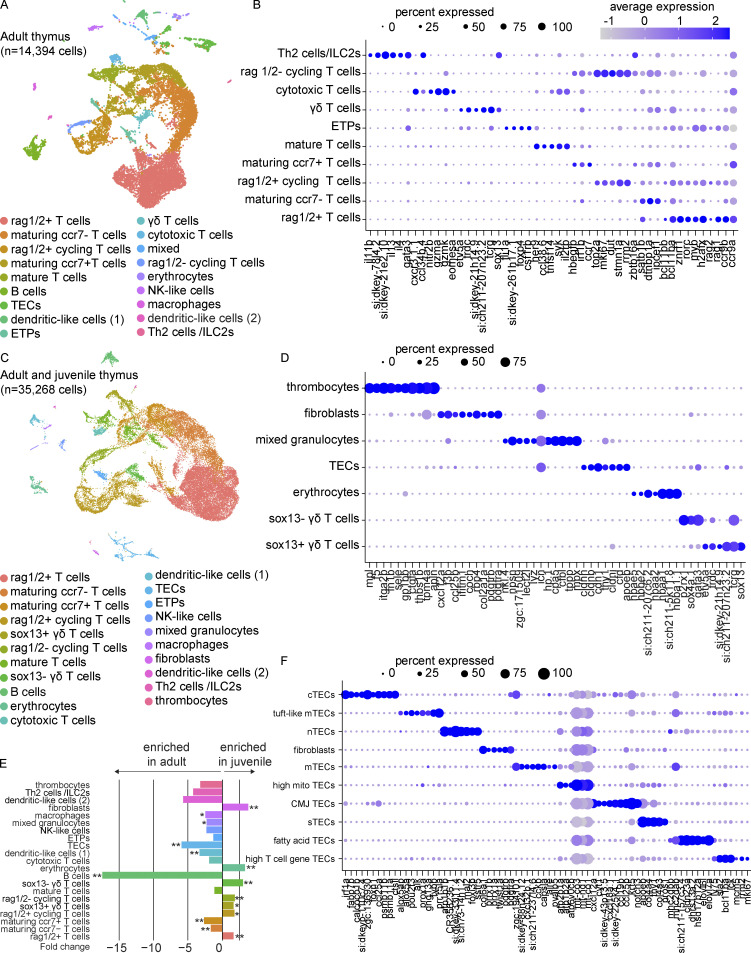
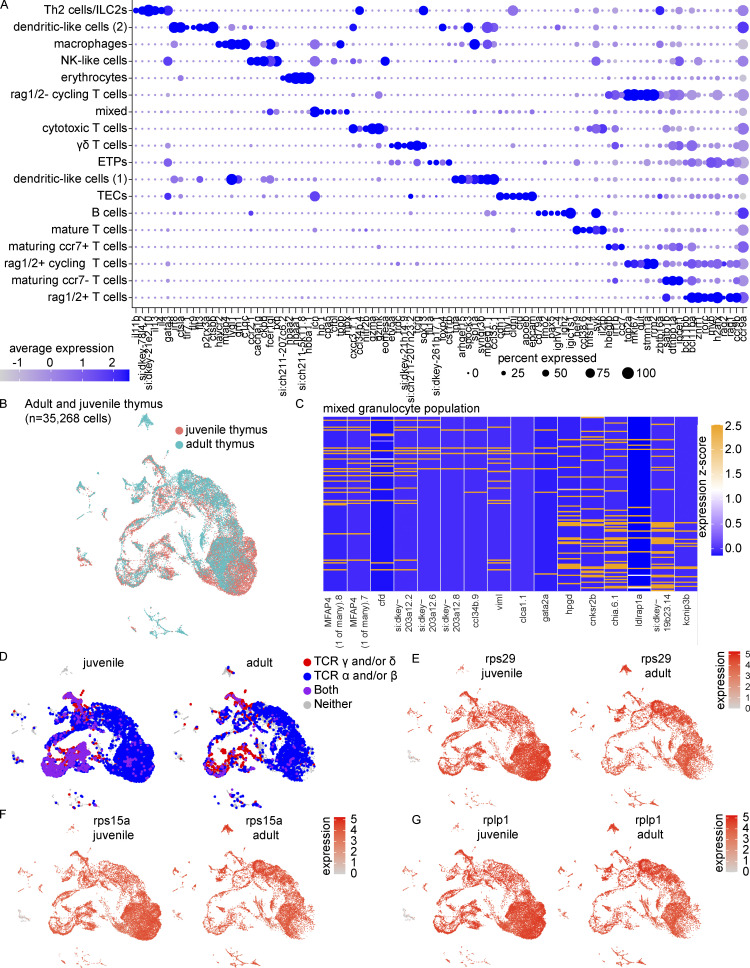
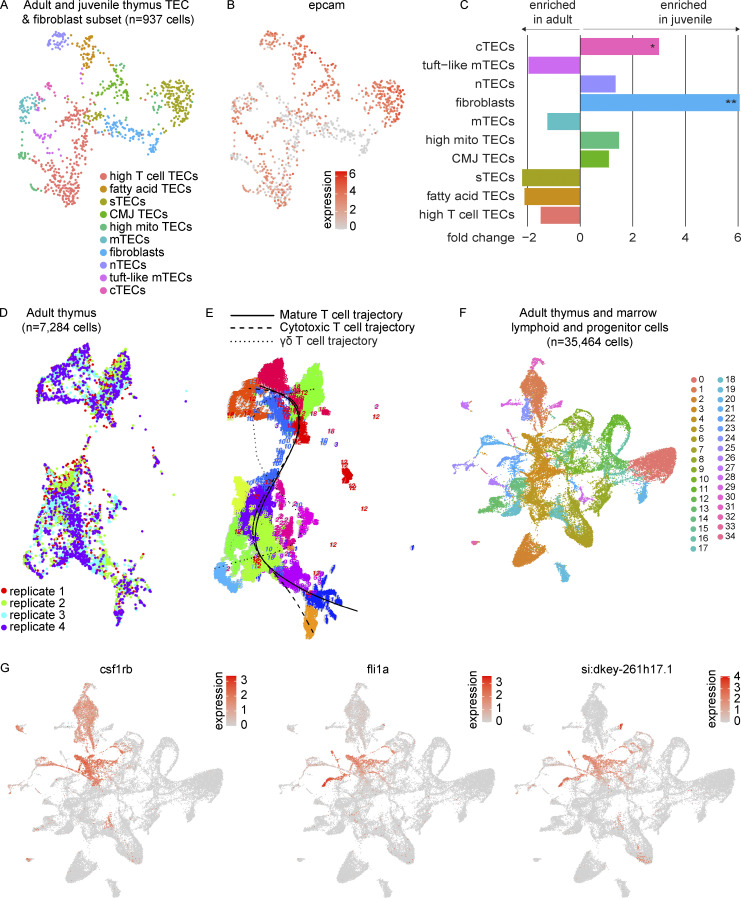
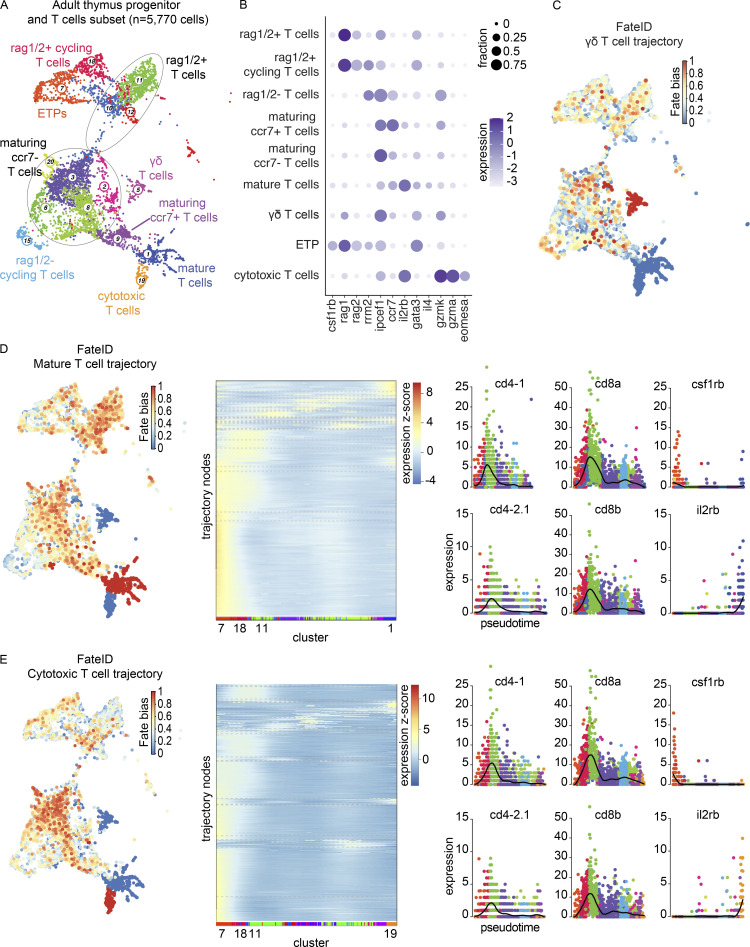
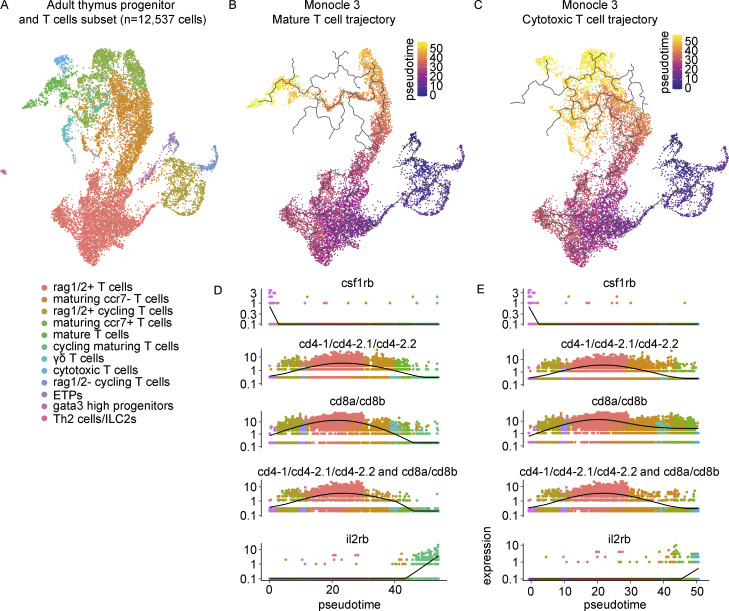
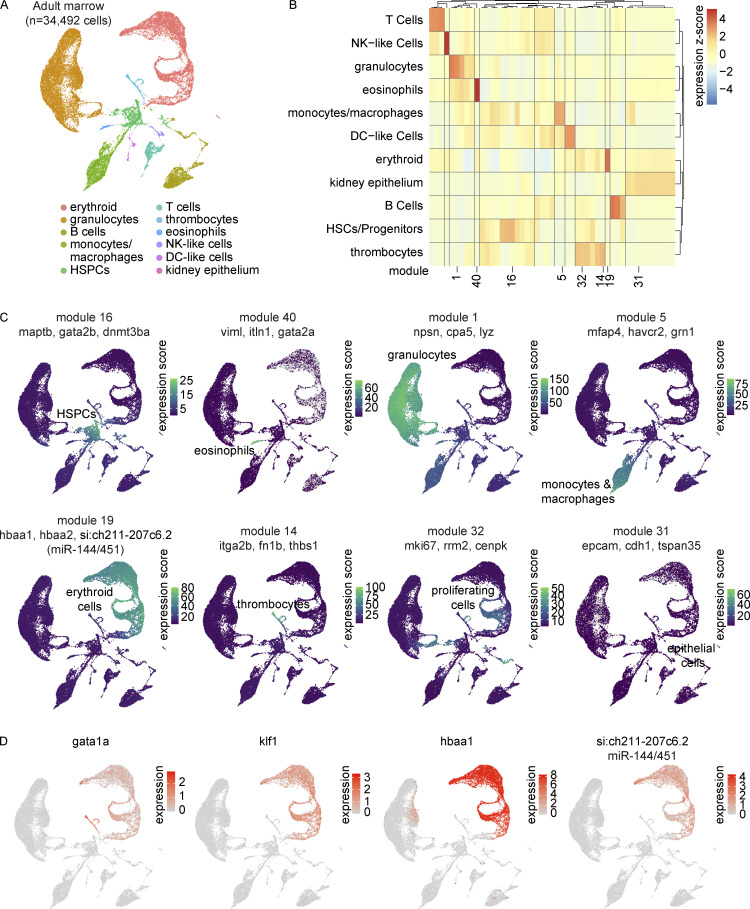
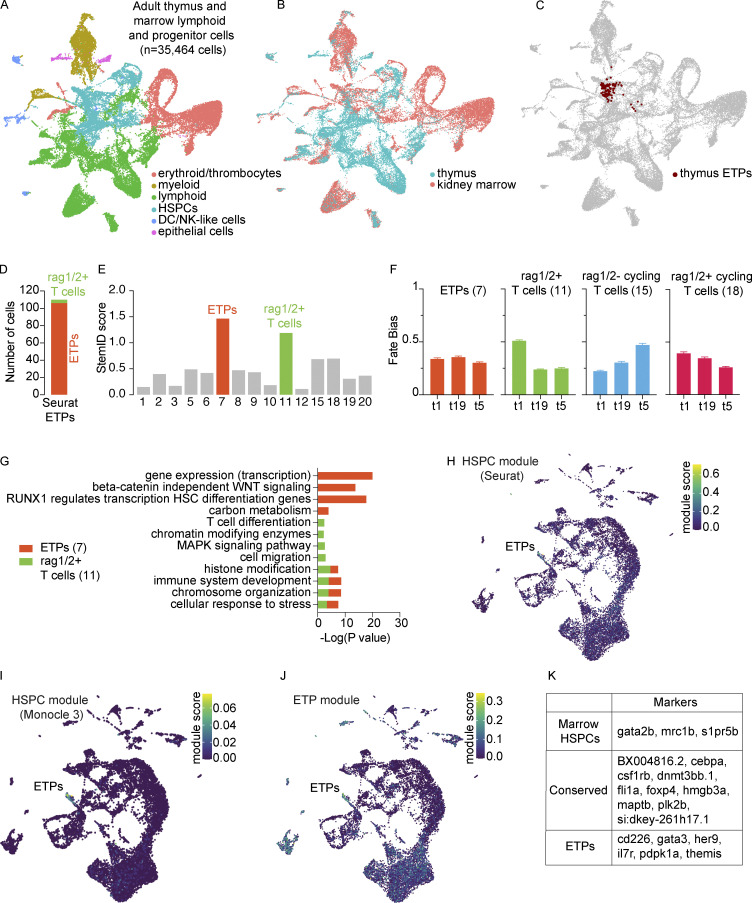
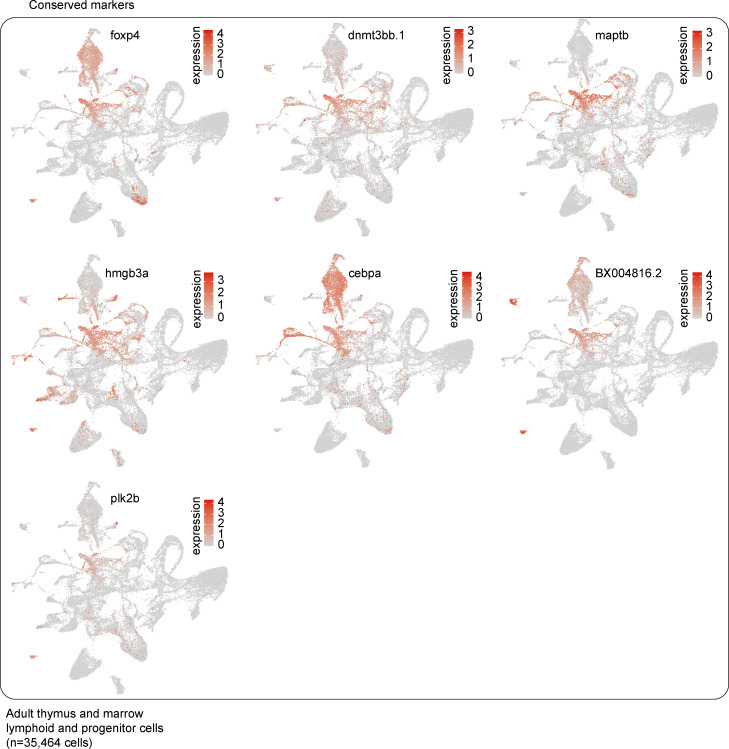
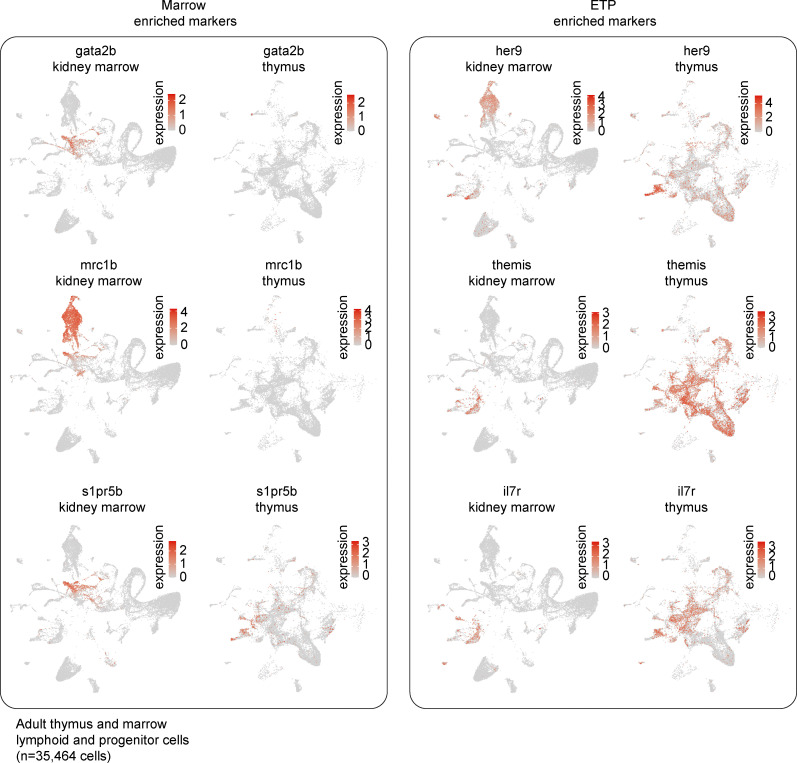
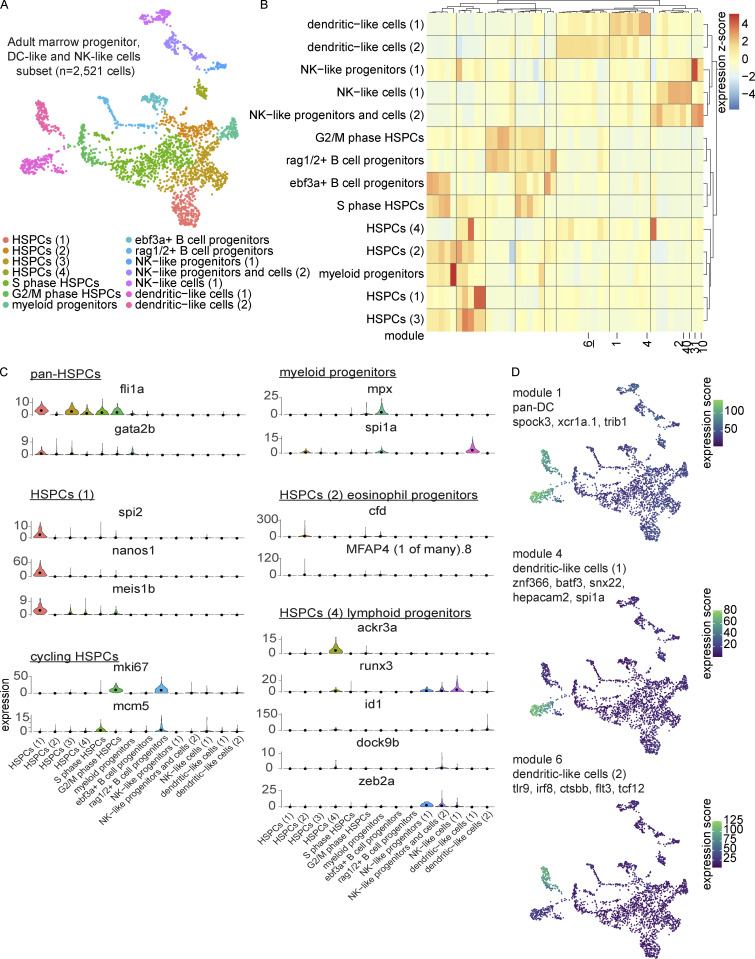
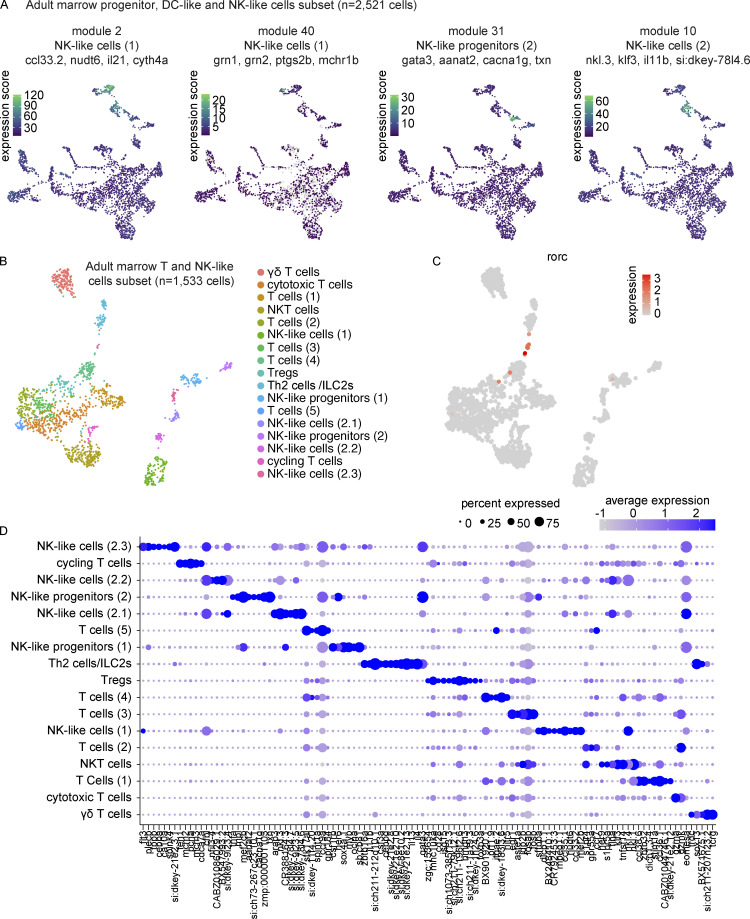
The zebrafish has proven to be a valuable model organism for studying hematopoiesis, but relatively little is known about zebrafish immune cell development and functional diversity. Elucidating key aspects of zebrafish lymphocyte development and exploring the breadth of effector functions would provide valuable insight into the evolution of adaptive immunity. We performed single-cell RNA sequencing on ∼70,000 cells from the zebrafish marrow and thymus to establish a gene expression map of zebrafish immune cell development. We uncovered rich cellular diversity in the juvenile and adult zebrafish thymus, elucidated B- and T-cell developmental trajectories, and transcriptionally characterized subsets of hematopoietic stem and progenitor cells and early thymic progenitors. Our analysis permitted the identification of two dendritic-like cell populations and provided evidence in support of the existence of a pre-B cell state. Our results provide critical insights into the landscape of zebrafish immunology and offer a foundation for cellular and genetic studies.
© 2022 Rubin et al.
Conflict of interest statement
Disclosures: L.I. Zon reported personal fees from Fate Therapeutics, CAMP4 Therapeutics, Amagma Therapeutics, Scholar Rock, Branch Biosciences, Celularity, and Cellarity outside the submitted work. No other disclosures were reported.
Figures
Similar articles
-
Single-Cell RNA Sequencing Resolves Spatiotemporal Development of Pre-thymic Lymphoid Progenitors and Thymus Organogenesis in Human Embryos.Immunity. 2019 Nov 19;51(5):930-948.e6. doi: 10.1016/j.immuni.2019.09.008. Epub 2019 Oct 8. Immunity. 2019. PMID: 31604687
-
Teleost growth factor independence (gfi) genes differentially regulate successive waves of hematopoiesis.Dev Biol. 2013 Jan 15;373(2):431-41. doi: 10.1016/j.ydbio.2012.08.015. Epub 2012 Aug 29. Dev Biol. 2013. PMID: 22960038 Free PMC article.
-
Fishing for lymphoid genes.Trends Immunol. 2001 Jun;22(6):302-7. doi: 10.1016/s1471-4906(01)01939-1. Trends Immunol. 2001. PMID: 11377288 Review.
-
Zebrafish model for allogeneic hematopoietic cell transplantation not requiring preconditioning.Proc Natl Acad Sci U S A. 2013 Mar 12;110(11):4327-32. doi: 10.1073/pnas.1219847110. Epub 2013 Feb 19. Proc Natl Acad Sci U S A. 2013. PMID: 23431192 Free PMC article.
-
Studying the adaptive immune system in zebrafish by transplantation of hematopoietic precursor cells.Methods Cell Biol. 2017;138:151-161. doi: 10.1016/bs.mcb.2016.08.003. Epub 2016 Sep 29. Methods Cell Biol. 2017. PMID: 28129842 Review.
Cited by
-
The axillary lymphoid organ - an external, experimentally accessible immune organ in the zebrafish.bioRxiv [Preprint]. 2024 Jul 25:2024.07.25.605139. doi: 10.1101/2024.07.25.605139. bioRxiv. 2024. PMID: 39091802 Free PMC article. Preprint.
-
Dynamic Changes in Lymphocyte Populations Establish Zebrafish as a Thymic Involution Model.J Immunol. 2024 Jun 1;212(11):1733-1743. doi: 10.4049/jimmunol.2300495. J Immunol. 2024. PMID: 38656392
-
Transcriptional Analysis of the Endostyle Reveals Pharyngeal Organ Functions in Ascidian.Biology (Basel). 2023 Feb 3;12(2):245. doi: 10.3390/biology12020245. Biology (Basel). 2023. PMID: 36829522 Free PMC article.
-
Evaluation of genetic demultiplexing of single-cell sequencing data from model species.Life Sci Alliance. 2023 May 17;6(8):e202301979. doi: 10.26508/lsa.202301979. Print 2023 Aug. Life Sci Alliance. 2023. PMID: 37197983 Free PMC article.
-
Catching the Big Fish in Big Data: A Meta-Analysis of Zebrafish Kidney scRNA-Seq Datasets Highlights Conserved Molecular Profiles of Macrophages and Neutrophils in Vertebrates.Biology (Basel). 2024 Sep 27;13(10):773. doi: 10.3390/biology13100773. Biology (Basel). 2024. PMID: 39452082 Free PMC article.
References
-
- Avagyan, S., Weber M.C., Ma S., Prasad M., Mannherz W.P., Yang S., Buenrostro J.D., and Zon L.I.. 2021. Single-cell ATAC-seq reveals GATA2-dependent priming defect in myeloid and a maturation bottleneck in lymphoid lineages. Blood Adv. 5:2673–2686. 10.1182/bloodadvances.2020002992 - DOI - PMC - PubMed
-
- Bachem, A., Hartung E., Guttler S., Mora A., Zhou X., Hegemann A., Plantinga M., Mazzini E., Stoitzner P., Gurka S., et al. . 2012. Expression of XCR1 characterizes the batf3-dependent lineage of dendritic cells capable of antigen cross-presentation. Front. Immunol. 3:214. 10.3389/fimmu.2012.00214 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
