

Live attenuated influenza A virus vaccines with modified NS1 proteins for veterinary use
- PMID: 35937688
- PMCID: PMC9354547
- DOI: 10.3389/fcimb.2022.954811
Live attenuated influenza A virus vaccines with modified NS1 proteins for veterinary use
Abstract
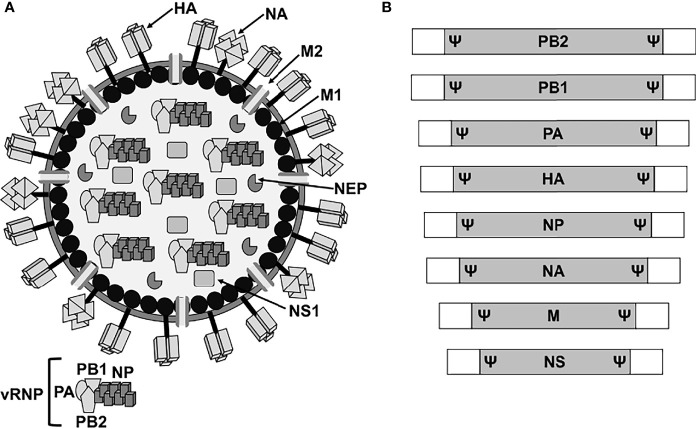
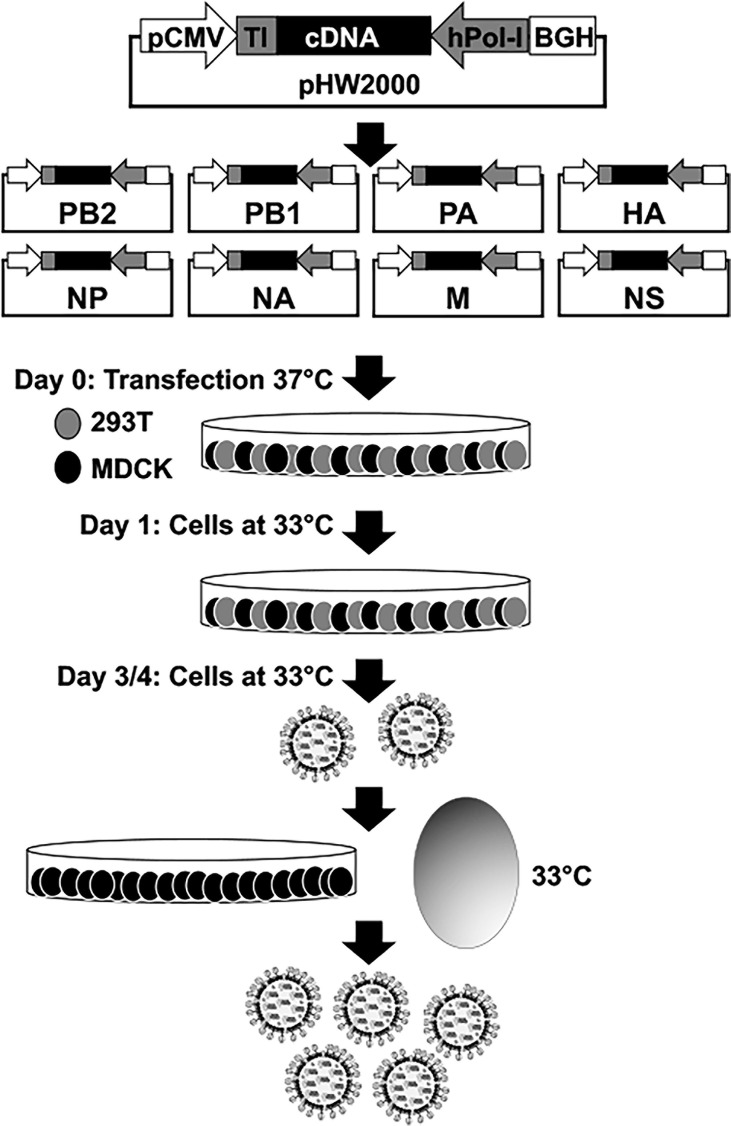
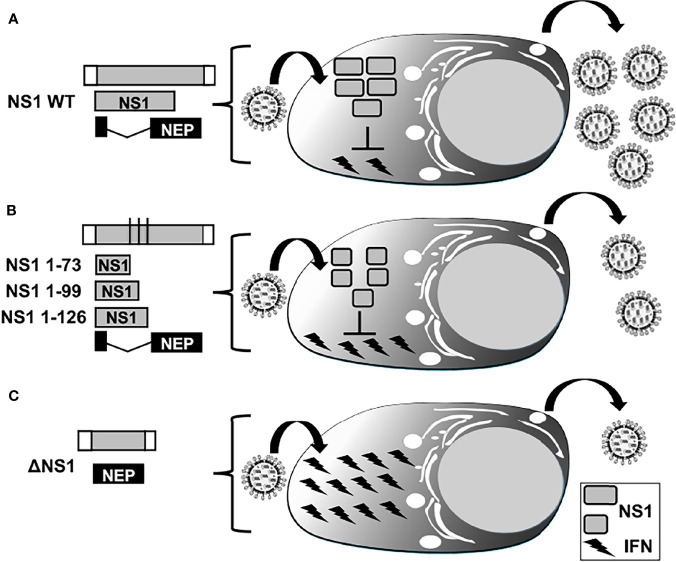
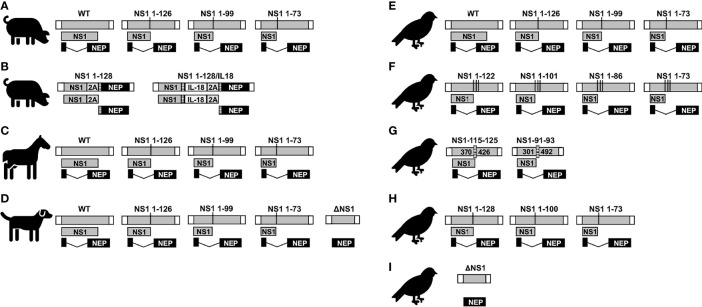
Influenza A viruses (IAV) spread rapidly and can infect a broad range of avian or mammalian species, having a tremendous impact in human and animal health and the global economy. IAV have evolved to develop efficient mechanisms to counteract innate immune responses, the first host mechanism that restricts IAV infection and replication. One key player in this fight against host-induced innate immune responses is the IAV non-structural 1 (NS1) protein that modulates antiviral responses and virus pathogenicity during infection. In the last decades, the implementation of reverse genetics approaches has allowed to modify the viral genome to design recombinant IAV, providing researchers a powerful platform to develop effective vaccine strategies. Among them, different levels of truncation or deletion of the NS1 protein of multiple IAV strains has resulted in attenuated viruses able to induce robust innate and adaptive immune responses, and high levels of protection against wild-type (WT) forms of IAV in multiple animal species and humans. Moreover, this strategy allows the development of novel assays to distinguish between vaccinated and/or infected animals, also known as Differentiating Infected from Vaccinated Animals (DIVA) strategy. In this review, we briefly discuss the potential of NS1 deficient or truncated IAV as safe, immunogenic and protective live-attenuated influenza vaccines (LAIV) to prevent disease caused by this important animal and human pathogen.
Keywords: differentiating infected from vaccinated animals (DIVA); influenza A virus; interferon; live-attenuated influenza vaccine (LAIV); non-structural 1 (NS1) protein.
Copyright © 2022 Nogales, DeDiego and Martínez-Sobrido.
Conflict of interest statement
LM-S and AN have patented LAIV for the prevention of canine and equine IAV. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins.Viruses. 2018 Dec 12;10(12):708. doi: 10.3390/v10120708. Viruses. 2018. PMID: 30545063 Free PMC article. Review.
-
Interplay of PA-X and NS1 Proteins in Replication and Pathogenesis of a Temperature-Sensitive 2009 Pandemic H1N1 Influenza A Virus.J Virol. 2017 Aug 10;91(17):e00720-17. doi: 10.1128/JVI.00720-17. Print 2017 Sep 1. J Virol. 2017. PMID: 28637750 Free PMC article.
-
Generation of DelNS1 Influenza Viruses: a Strategy for Optimizing Live Attenuated Influenza Vaccines.mBio. 2019 Sep 17;10(5):e02180-19. doi: 10.1128/mBio.02180-19. mBio. 2019. PMID: 31530680 Free PMC article.
-
Differential Modulation of Innate Immune Responses in Human Primary Cells by Influenza A Viruses Carrying Human or Avian Nonstructural Protein 1.J Virol. 2019 Dec 12;94(1):e00999-19. doi: 10.1128/JVI.00999-19. Print 2019 Dec 12. J Virol. 2019. PMID: 31597767 Free PMC article.
-
Attenuated influenza virus vaccines with modified NS1 proteins.Curr Top Microbiol Immunol. 2009;333:177-95. doi: 10.1007/978-3-540-92165-3_9. Curr Top Microbiol Immunol. 2009. PMID: 19768406 Review.
Cited by
-
Bacterial Artificial Chromosome Reverse Genetics Approaches for SARS-CoV-2.Methods Mol Biol. 2024;2733:133-153. doi: 10.1007/978-1-0716-3533-9_9. Methods Mol Biol. 2024. PMID: 38064031
-
Recombinant Influenza A Viruses Expressing Reporter Genes from the Viral NS Segment.Int J Mol Sci. 2024 Oct 1;25(19):10584. doi: 10.3390/ijms251910584. Int J Mol Sci. 2024. PMID: 39408912 Free PMC article. Review.
-
Reprogramming viral immune evasion for a rational design of next-generation vaccines for RNA viruses.Front Immunol. 2023 Apr 17;14:1172000. doi: 10.3389/fimmu.2023.1172000. eCollection 2023. Front Immunol. 2023. PMID: 37138878 Free PMC article. Review.
-
FHL1 promotes chikungunya and o'nyong-nyong virus infection and pathogenesis with implications for alphavirus vaccine design.Nat Commun. 2023 Oct 26;14(1):6605. doi: 10.1038/s41467-023-42330-2. Nat Commun. 2023. PMID: 37884534 Free PMC article.
-
Mass vaccination with reassortment-impaired live H9N2 avian influenza vaccine.NPJ Vaccines. 2024 Aug 3;9(1):136. doi: 10.1038/s41541-024-00923-y. NPJ Vaccines. 2024. PMID: 39097573 Free PMC article.
References
-
- (2015). Outbreak of canine influenza caused by new strain of virus. Javma-Journal Am. Veterinary Med. Assoc. 246 (10), 1049–1049.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical