

Topological digestion drives time-varying rheology of entangled DNA fluids
- PMID: 35902575
- PMCID: PMC9334285
- DOI: 10.1038/s41467-022-31828-w
Topological digestion drives time-varying rheology of entangled DNA fluids
Abstract
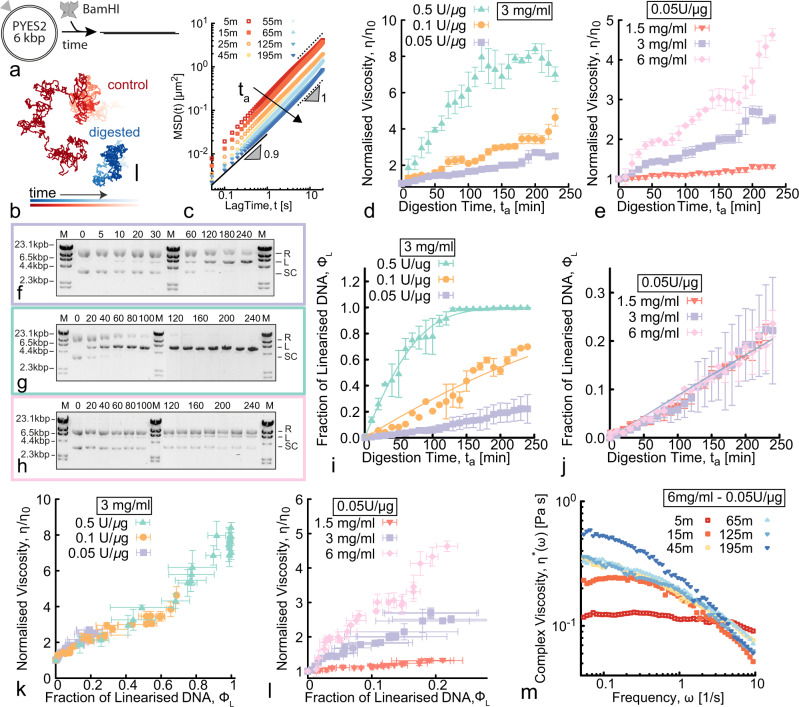
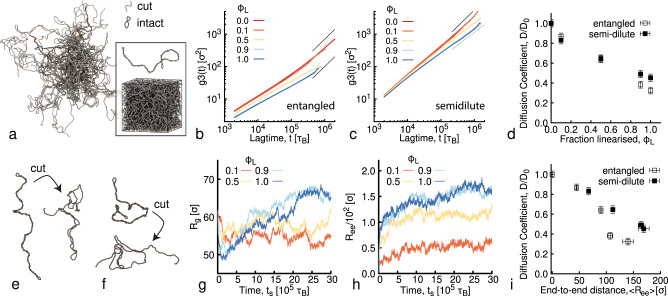
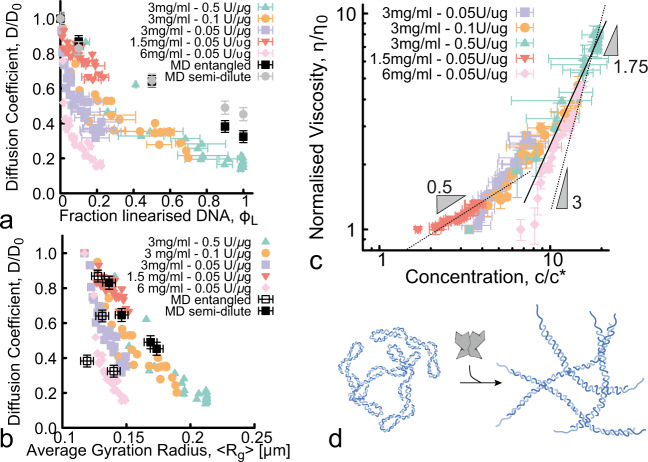
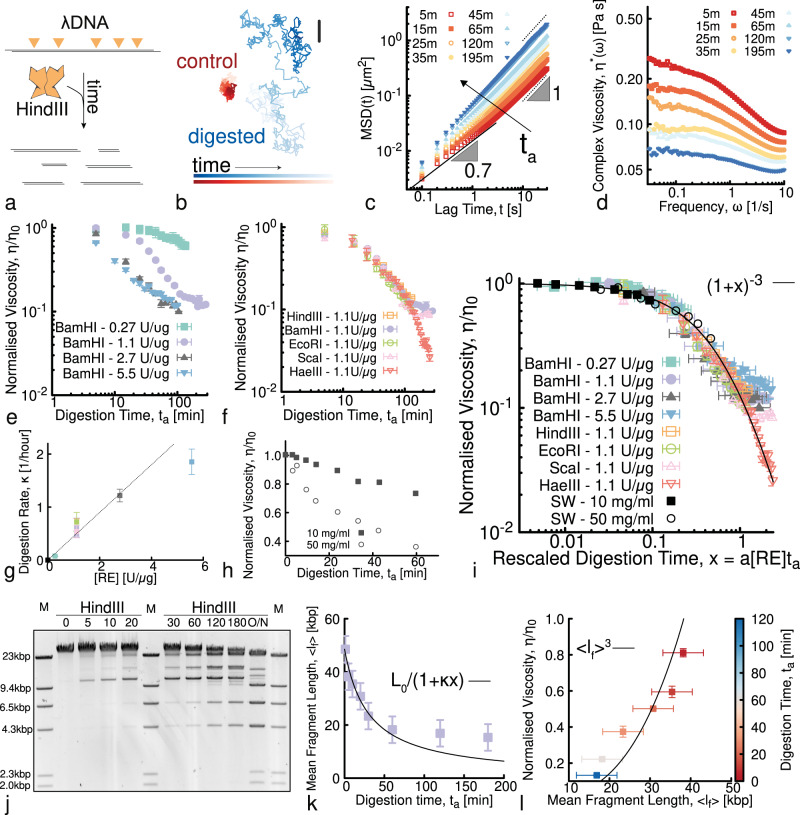
Understanding and controlling the rheology of polymeric complex fluids that are pushed out-of-equilibrium is a fundamental problem in both industry and biology. For example, to package, repair, and replicate DNA, cells use enzymes to constantly manipulate DNA topology, length, and structure. Inspired by this feat, here we engineer and study DNA-based complex fluids that undergo enzymatically-driven topological and architectural alterations via restriction endonuclease (RE) reactions. We show that these systems display time-dependent rheological properties that depend on the concentrations and properties of the comprising DNA and REs. Through time-resolved microrheology experiments and Brownian Dynamics simulations, we show that conversion of supercoiled to linear DNA topology leads to a monotonic increase in viscosity. On the other hand, the viscosity of entangled linear DNA undergoing fragmentation displays a universal decrease that we rationalise using living polymer theory. Finally, to showcase the tunability of these behaviours, we design a DNA fluid that exhibits a time-dependent increase, followed by a temporally-gated decrease, of its viscosity. Our results present a class of polymeric fluids that leverage naturally occurring enzymes to drive diverse time-varying rheology by performing architectural alterations to the constituents.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Cooperative Rheological State-Switching of Enzymatically-Driven Composites of Circular DNA And Dextran.Adv Mater. 2023 Nov;35(46):e2305824. doi: 10.1002/adma.202305824. Epub 2023 Oct 15. Adv Mater. 2023. PMID: 37500570
-
Enzymatic cleaving of entangled DNA rings drives scale-dependent rheological trajectories.Soft Matter. 2024 Mar 20;20(12):2750-2766. doi: 10.1039/d3sm01641b. Soft Matter. 2024. PMID: 38440846
-
Unexpected entanglement dynamics in semidilute blends of supercoiled and ring DNA.Soft Matter. 2020 Jan 7;16(1):152-161. doi: 10.1039/c9sm01767d. Epub 2019 Nov 27. Soft Matter. 2020. PMID: 31774103
-
DNA as a Model for Probing Polymer Entanglements: Circular Polymers and Non-Classical Dynamics.Polymers (Basel). 2016 Sep 7;8(9):336. doi: 10.3390/polym8090336. Polymers (Basel). 2016. PMID: 30974610 Free PMC article. Review.
-
Advances and challenges in the high-pressure rheology of complex fluids.Adv Colloid Interface Sci. 2021 Aug;294:102472. doi: 10.1016/j.cis.2021.102472. Epub 2021 Jun 26. Adv Colloid Interface Sci. 2021. PMID: 34311156 Review.
Cited by
-
Worm blobs as entangled living polymers: from topological active matter to flexible soft robot collectives.Soft Matter. 2023 Sep 27;19(37):7057-7069. doi: 10.1039/d3sm00542a. Soft Matter. 2023. PMID: 37706563 Free PMC article. Review.
-
Topological Considerations in Biomolecular Condensation.Biomolecules. 2023 Jan 11;13(1):151. doi: 10.3390/biom13010151. Biomolecules. 2023. PMID: 36671536 Free PMC article. Review.
-
Effects of monovalent and divalent cations on the rheology of entangled DNA.Soft Matter. 2024 May 15;20(19):3980-3986. doi: 10.1039/d3sm00957b. Soft Matter. 2024. PMID: 38686506 Free PMC article.
References
-
- Calladine, C. R., Drew, H. R., Luisi, B. F. & Travers, A. A. Understanding DNA: the Molecule and How It Works. 3rd edn (Elsevier Academic Press, 2004).
-
- Bates, A. & Maxwell, A. DNA Topology (Oxford University Press, 2005).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
