

The Secreted Ribonuclease SRE1 Contributes to Setosphaeria turcica Virulence and Activates Plant Immunity
- PMID: 35875548
- PMCID: PMC9304870
- DOI: 10.3389/fmicb.2022.941991
The Secreted Ribonuclease SRE1 Contributes to Setosphaeria turcica Virulence and Activates Plant Immunity
Abstract
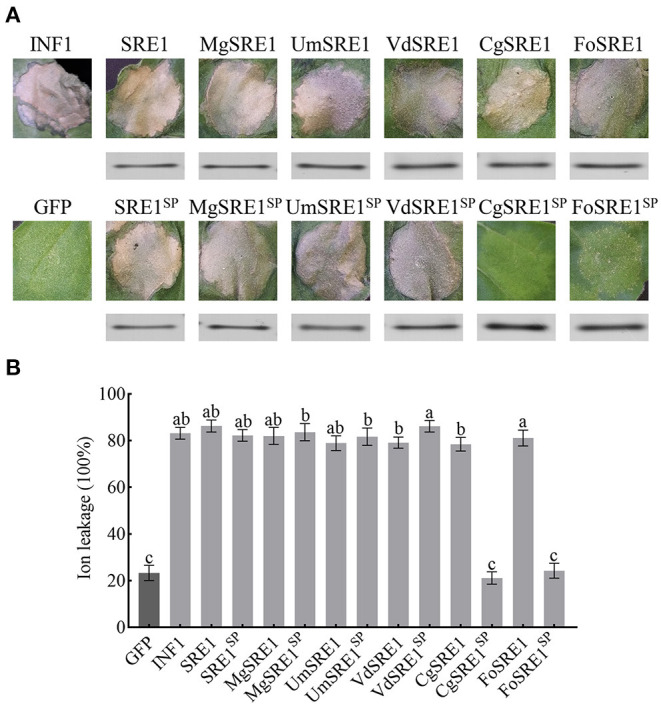
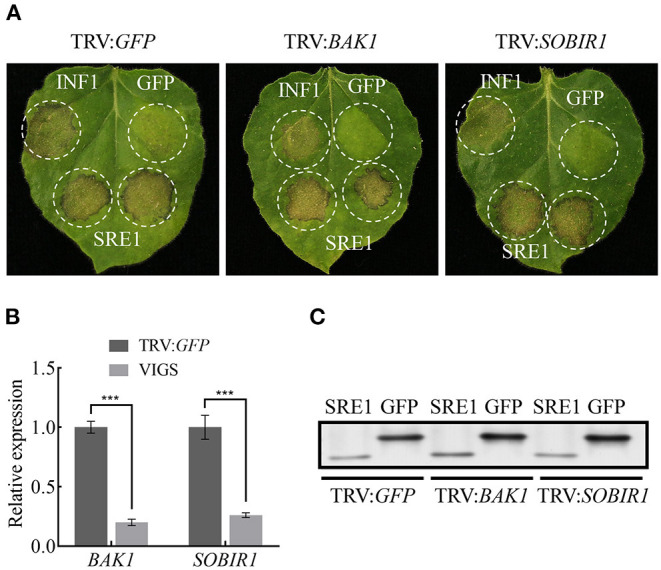
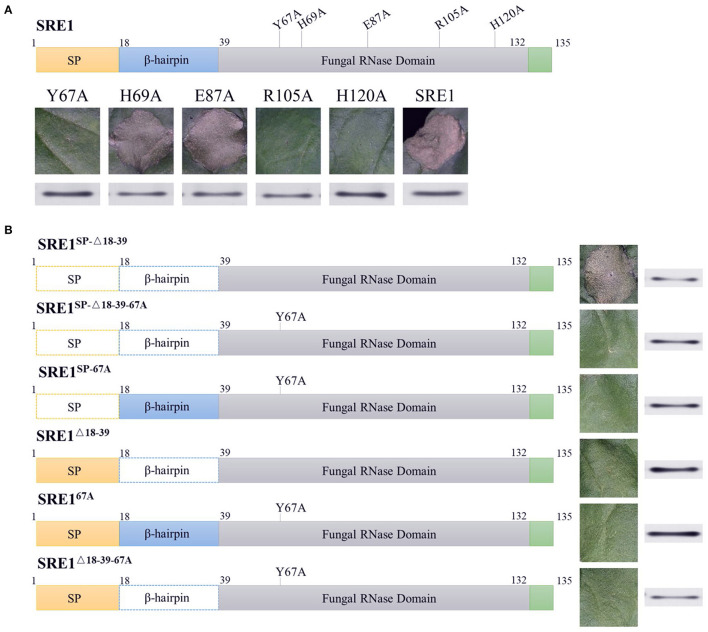
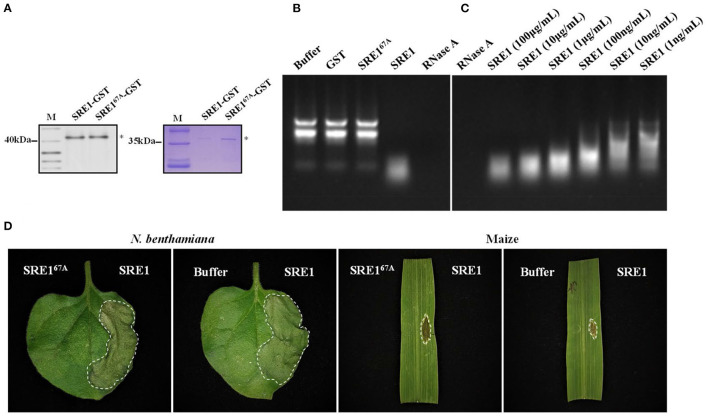
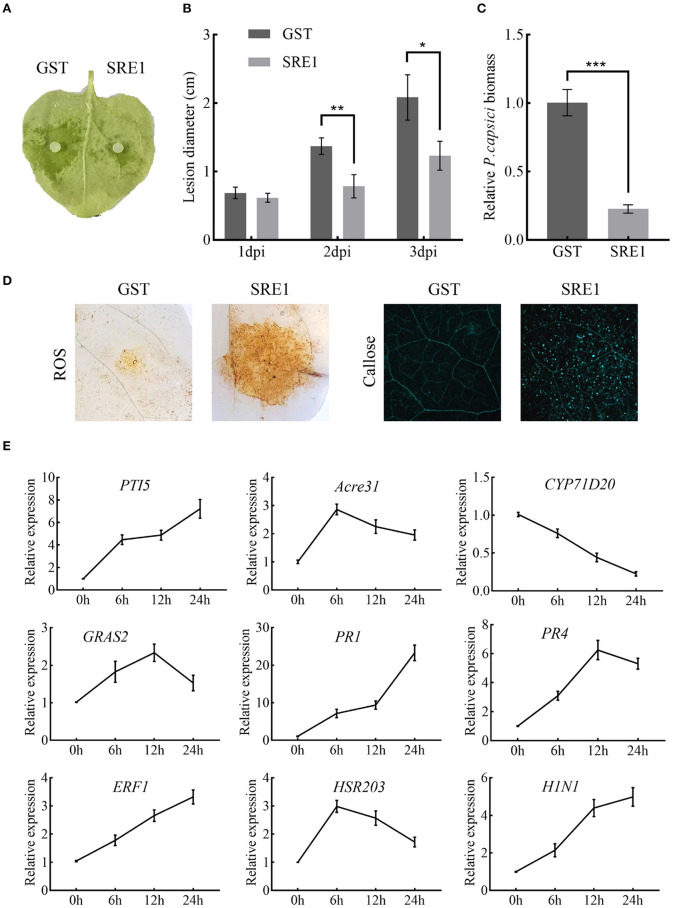
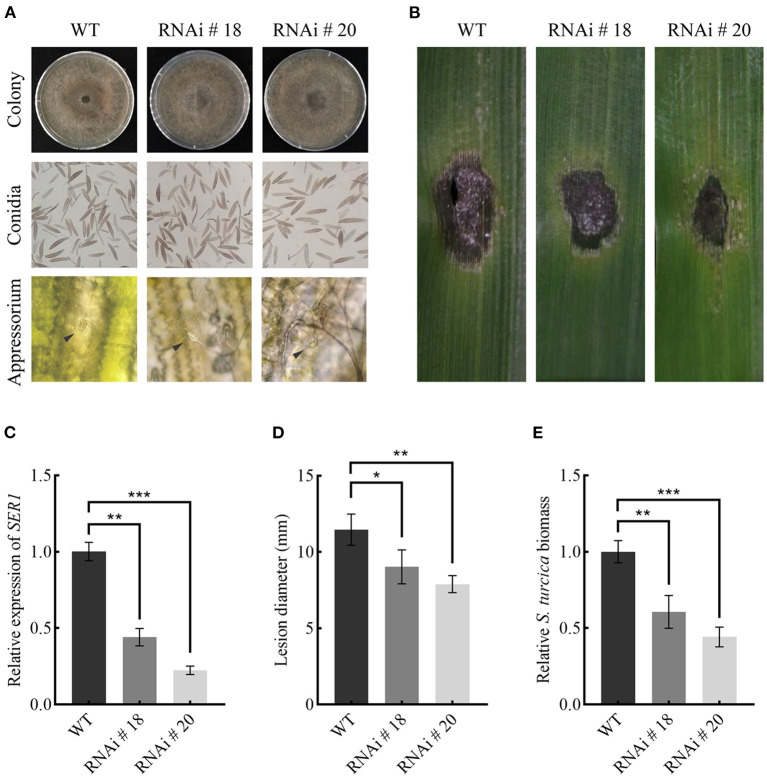
During the plant infection process, pathogens can secrete several effectors. Some of the effectors are well-known for their roles in regulating plant immunity and promoting successful pathogen colonization. However, there are few studies on the ribonuclease (RNase) effectors secreted by fungi. In the present study, we discovered a secretable RNase (SRE1) in the secretome of Setosphaeria turcica that was significantly upregulated during the early stages of S. turcica infection in maize. Knockdown of SRE1 significantly reduced the virulence of S. turcica. SRE1 can induce cell death in maize and Nicotiana benthamiana. However, unlike the conventional hypersensitive response (HR) caused by other effectors, SRE1 is not dependent on its signal peptide (SP) or plant receptor kinases (such as BAK1 and SOBIR1). SRE1-induced cell death depends upon its enzymatic activity and the N-terminal β-hairpin structure. SRE1 relies on its N-terminal β-hairpin structure to enter cells, and then degrades plant's RNA through its catalytic activity causing cytotoxic effects. Additionally, SRE1 enhances N. benthamiana's resistance to pathogenic fungi and oomycetes. In summary, SRE1 promotes the virulence of S. turcica, inducing plant cell death and activating plant immune responses.
Keywords: Setosphaeria turcica; cell death; maize; plant immunity; ribonuclease.
Copyright © 2022 He, Huang, Sun, Liu, Wang, Xuan and Gao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Fg12 ribonuclease secretion contributes to Fusarium graminearum virulence and induces plant cell death.J Integr Plant Biol. 2021 Feb;63(2):365-377. doi: 10.1111/jipb.12997. Epub 2020 Aug 22. J Integr Plant Biol. 2021. PMID: 32725938
-
Novel factors contributing to fungal pathogenicity at early stages of Setosphaeria turcica infection.Mol Plant Pathol. 2022 Jan;23(1):32-44. doi: 10.1111/mpp.13140. Epub 2021 Oct 10. Mol Plant Pathol. 2022. PMID: 34628700 Free PMC article.
-
Setosphaeria rostrata: Insights from the sequenced genome of Setosphaeria turcica.Fungal Genet Biol. 2013 Dec;61:158-63. doi: 10.1016/j.fgb.2013.08.015. Epub 2013 Sep 5. Fungal Genet Biol. 2013. PMID: 24012822
-
Microbial Effectors: Key Determinants in Plant Health and Disease.Microorganisms. 2022 Oct 6;10(10):1980. doi: 10.3390/microorganisms10101980. Microorganisms. 2022. PMID: 36296254 Free PMC article. Review.
-
Versatile effectors of phytopathogenic fungi target host immunity.J Integr Plant Biol. 2021 Nov;63(11):1856-1873. doi: 10.1111/jipb.13162. Epub 2021 Sep 15. J Integr Plant Biol. 2021. PMID: 34383388 Review.
Cited by
-
Structural polymorphisms within a common powdery mildew effector scaffold as a driver of coevolution with cereal immune receptors.Proc Natl Acad Sci U S A. 2023 Aug 8;120(32):e2307604120. doi: 10.1073/pnas.2307604120. Epub 2023 Jul 31. Proc Natl Acad Sci U S A. 2023. PMID: 37523523 Free PMC article.
References
LinkOut - more resources
Full Text Sources