

Control of animal virus replication by RNA adenosine methylation
- PMID: 35840182
- PMCID: PMC10259266
- DOI: 10.1016/bs.aivir.2022.01.002
Control of animal virus replication by RNA adenosine methylation
Abstract
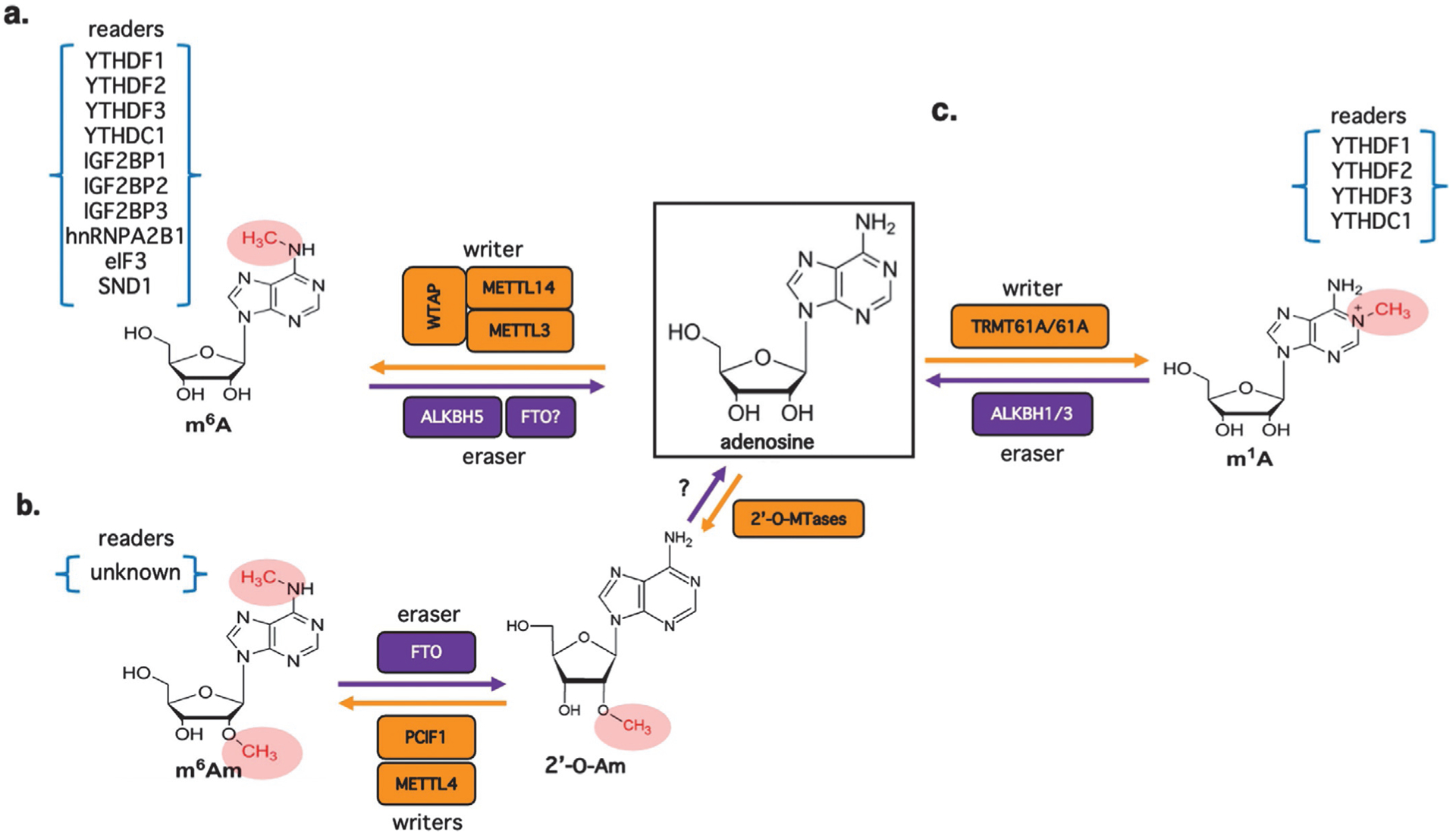
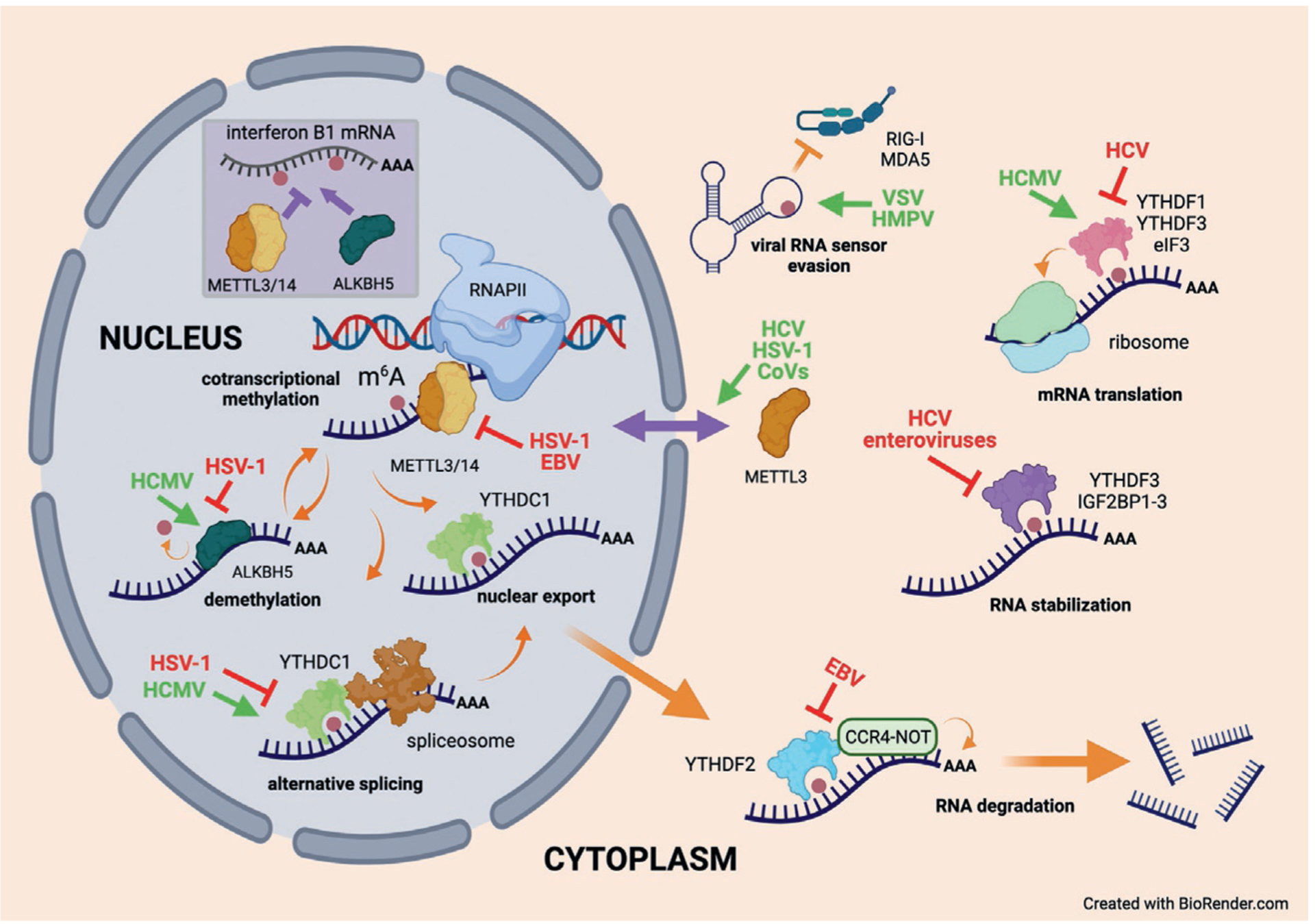
Methylation at the N6-position of either adenosine (m6A) or 2'-O-methyladenosine (m6Am) represents two of the most abundant internal modifications of coding and non-coding RNAs, influencing their maturation, stability and function. Additionally, although less abundant and less well-studied, monomethylation at the N1-position (m1A) can have profound effects on RNA folding. It has been known for several decades that RNAs produced by both DNA and RNA viruses can be m6A/m6Am modified and the list continues to broaden through advances in detection technologies and identification of the relevant methyltransferases. Recent studies have uncovered varied mechanisms used by viruses to manipulate the m6A pathway in particular, either to enhance virus replication or to antagonize host antiviral defenses. As such, RNA modifications represent an important frontier of exploration in the broader realm of virus-host interactions, and this new knowledge already suggests exciting opportunities for therapeutic intervention. In this review we summarize the principal mechanisms by which m6A/m6Am can promote or hinder viral replication, describe how the pathway is actively manipulated by biomedically important viruses, and highlight some remaining gaps in understanding how adenosine methylation of RNA controls viral replication and pathogenesis.
Keywords: Epitranscriptomic regulation; N(1)-methyladenosine; N(6),2′-O-methyladenosine; N(6)-methyladenosine; RNA modification; Virus–host interactions.
Copyright © 2022 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Nonsegmented Negative-Sense RNA Viruses Utilize N6-Methyladenosine (m6A) as a Common Strategy To Evade Host Innate Immunity.J Virol. 2021 Apr 12;95(9):e01939-20. doi: 10.1128/JVI.01939-20. Print 2021 Apr 12. J Virol. 2021. PMID: 33536170 Free PMC article.
-
Epitranscriptomic(N6-methyladenosine) Modification of Viral RNA and Virus-Host Interactions.Front Cell Infect Microbiol. 2020 Nov 24;10:584283. doi: 10.3389/fcimb.2020.584283. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33330128 Free PMC article. Review.
-
N6-methyladenosine regulates PEDV replication and host gene expression.Virology. 2020 Sep;548:59-72. doi: 10.1016/j.virol.2020.06.008. Epub 2020 Jun 16. Virology. 2020. PMID: 32838947 Free PMC article.
-
2'-O methylation of internal adenosine by flavivirus NS5 methyltransferase.PLoS Pathog. 2012;8(4):e1002642. doi: 10.1371/journal.ppat.1002642. Epub 2012 Apr 5. PLoS Pathog. 2012. PMID: 22496660 Free PMC article.
-
Regulatory effect of m6 A modification on different viruses.J Med Virol. 2021 Nov;93(11):6100-6115. doi: 10.1002/jmv.27246. Epub 2021 Aug 16. J Med Virol. 2021. PMID: 34329499 Review.
Cited by
-
1mΨ influences the performance of various positive-stranded RNA virus-based replicons.Sci Rep. 2024 Jul 31;14(1):17634. doi: 10.1038/s41598-024-68617-y. Sci Rep. 2024. PMID: 39085360 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
