

Leishmania guyanensis M4147 as a new LRV1-bearing model parasite: Phosphatidate phosphatase 2-like protein controls cell cycle progression and intracellular lipid content
- PMID: 35749562
- PMCID: PMC9232130
- DOI: 10.1371/journal.pntd.0010510
Leishmania guyanensis M4147 as a new LRV1-bearing model parasite: Phosphatidate phosphatase 2-like protein controls cell cycle progression and intracellular lipid content
Abstract
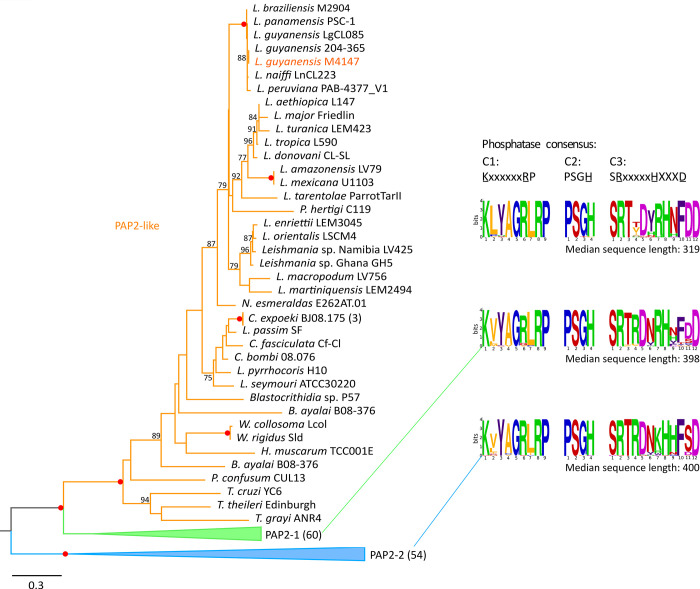
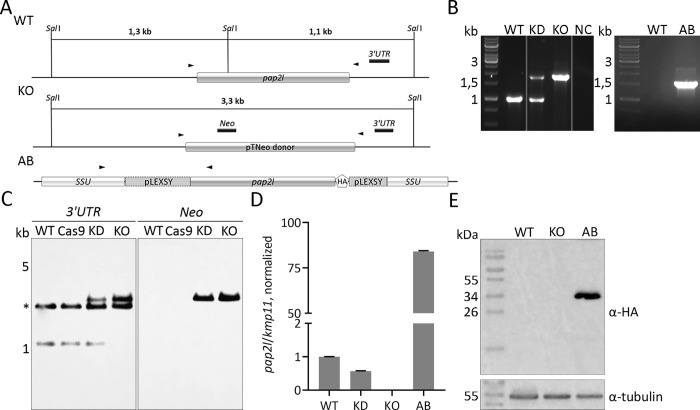
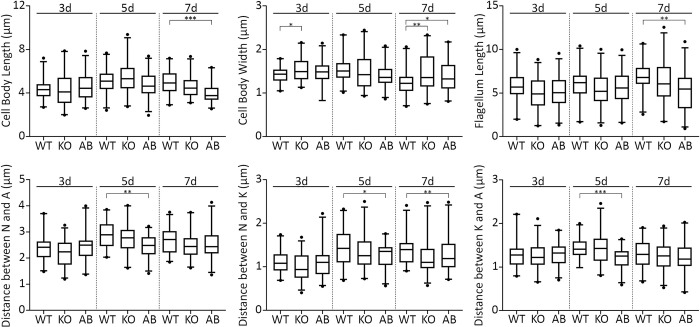
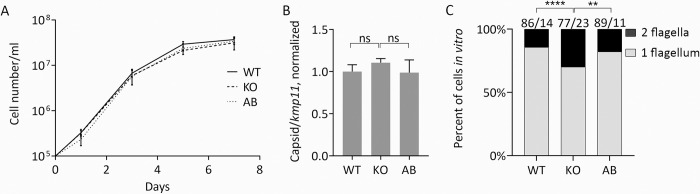
Leishmaniasis is a parasitic vector-borne disease caused by the protistan flagellates of the genus Leishmania. Leishmania (Viannia) guyanensis is one of the most common causative agents of the American tegumentary leishmaniasis. It has previously been shown that L. guyanensis strains that carry the endosymbiotic Leishmania RNA virus 1 (LRV1) cause more severe form of the disease in a mouse model than those that do not. The presence of the virus was implicated into the parasite's replication and spreading. In this respect, studying the molecular mechanisms of cellular control of viral infection is of great medical importance. Here, we report ~30.5 Mb high-quality genome assembly of the LRV1-positive L. guyanensis M4147. This strain was turned into a model by establishing the CRISPR-Cas9 system and ablating the gene encoding phosphatidate phosphatase 2-like (PAP2L) protein. The orthologue of this gene is conspicuously absent from the genome of an unusual member of the family Trypanosomatidae, Vickermania ingenoplastis, a species with mostly bi-flagellated cells. Our analysis of the PAP2L-null L. guyanensis showed an increase in the number of cells strikingly resembling the bi-flagellated V. ingenoplastis, likely as a result of the disruption of the cell cycle, significant accumulation of phosphatidic acid, and increased virulence compared to the wild type cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Presence of Leishmania RNA Virus 1 in Leishmania guyanensis Increases the Risk of First-Line Treatment Failure and Symptomatic Relapse.J Infect Dis. 2016 Jan 1;213(1):105-11. doi: 10.1093/infdis/jiv355. Epub 2015 Jun 29. J Infect Dis. 2016. PMID: 26123564
-
Leishmania (Viannia) guyanensis in tegumentary leishmaniasis.Pathog Dis. 2018 Jun 1;76(4). doi: 10.1093/femspd/fty025. Pathog Dis. 2018. PMID: 29722820 Review.
-
Leishmania Viannia guyanensis, LRV1 virus and extracellular vesicles: a dangerous trio influencing the faith of immune response during muco-cutaneous leishmaniasis.Curr Opin Immunol. 2020 Oct;66:108-113. doi: 10.1016/j.coi.2020.08.004. Epub 2020 Aug 30. Curr Opin Immunol. 2020. PMID: 32877837 Review.
-
Detection of Leishmania RNA Virus 1 in Leishmania (Viannia) panamensis Isolates, Panama.Emerg Infect Dis. 2023 Jun;29(6):1250-1253. doi: 10.3201/eid2906.220012. Emerg Infect Dis. 2023. PMID: 37209675 Free PMC article.
-
Raising the suspicion of a non-autochthonous infection: identification of Leishmania guyanensis from Costa Rica exhibits a Leishmaniavirus related to Brazilian north-east and French Guiana viral genotypes.Mem Inst Oswaldo Cruz. 2023 Jan 16;117:e220162. doi: 10.1590/0074-02760220162. eCollection 2023. Mem Inst Oswaldo Cruz. 2023. PMID: 36651455 Free PMC article.
Cited by
-
Impaired signaling pathways on Berardinelli-Seip congenital lipodystrophy macrophages during Leishmania infantum infection.Sci Rep. 2024 May 16;14(1):11236. doi: 10.1038/s41598-024-61663-6. Sci Rep. 2024. PMID: 38755198 Free PMC article.
-
TriTrypDB: An integrated functional genomics resource for kinetoplastida.PLoS Negl Trop Dis. 2023 Jan 19;17(1):e0011058. doi: 10.1371/journal.pntd.0011058. eCollection 2023 Jan. PLoS Negl Trop Dis. 2023. PMID: 36656904 Free PMC article.
-
Revisiting epidemiology of leishmaniasis in central Asia: lessons learnt.Parasitology. 2023 Feb;150(2):129-136. doi: 10.1017/S0031182022001640. Epub 2022 Dec 1. Parasitology. 2023. PMID: 36453145 Free PMC article. Review.
-
Elimination of LRVs Elicits Different Responses in Leishmania spp.mSphere. 2022 Aug 31;7(4):e0033522. doi: 10.1128/msphere.00335-22. Epub 2022 Aug 9. mSphere. 2022. PMID: 35943162 Free PMC article.
-
Evolution of RNA viruses in trypanosomatids: new insights from the analysis of Sauroleishmania.Parasitol Res. 2023 Oct;122(10):2279-2286. doi: 10.1007/s00436-023-07928-x. Epub 2023 Jul 25. Parasitol Res. 2023. PMID: 37490143 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
