

Context-specific Polycomb mechanisms in development
- PMID: 35681061
- PMCID: PMC9933872
- DOI: 10.1038/s41576-022-00499-0
Context-specific Polycomb mechanisms in development
Abstract
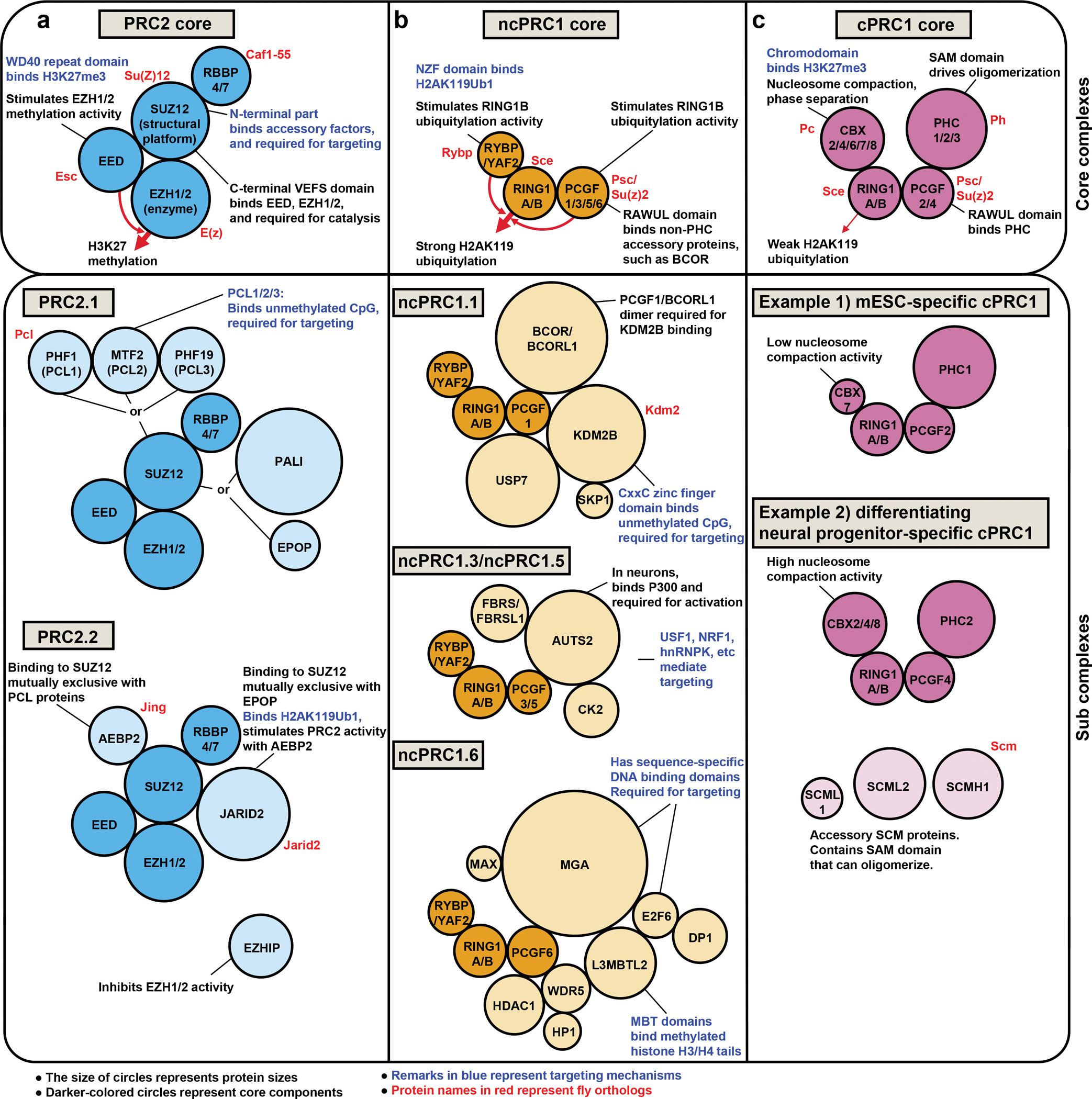
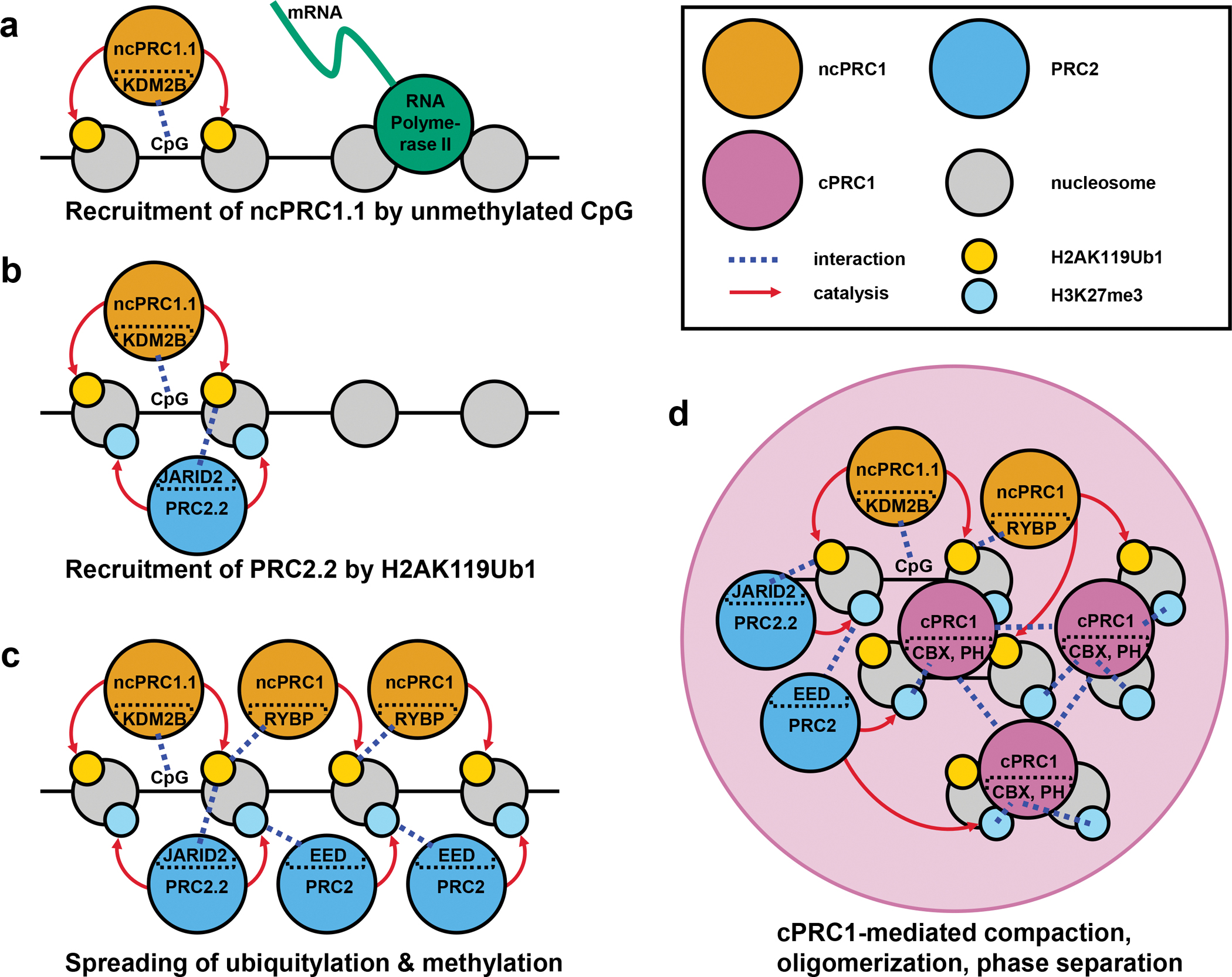
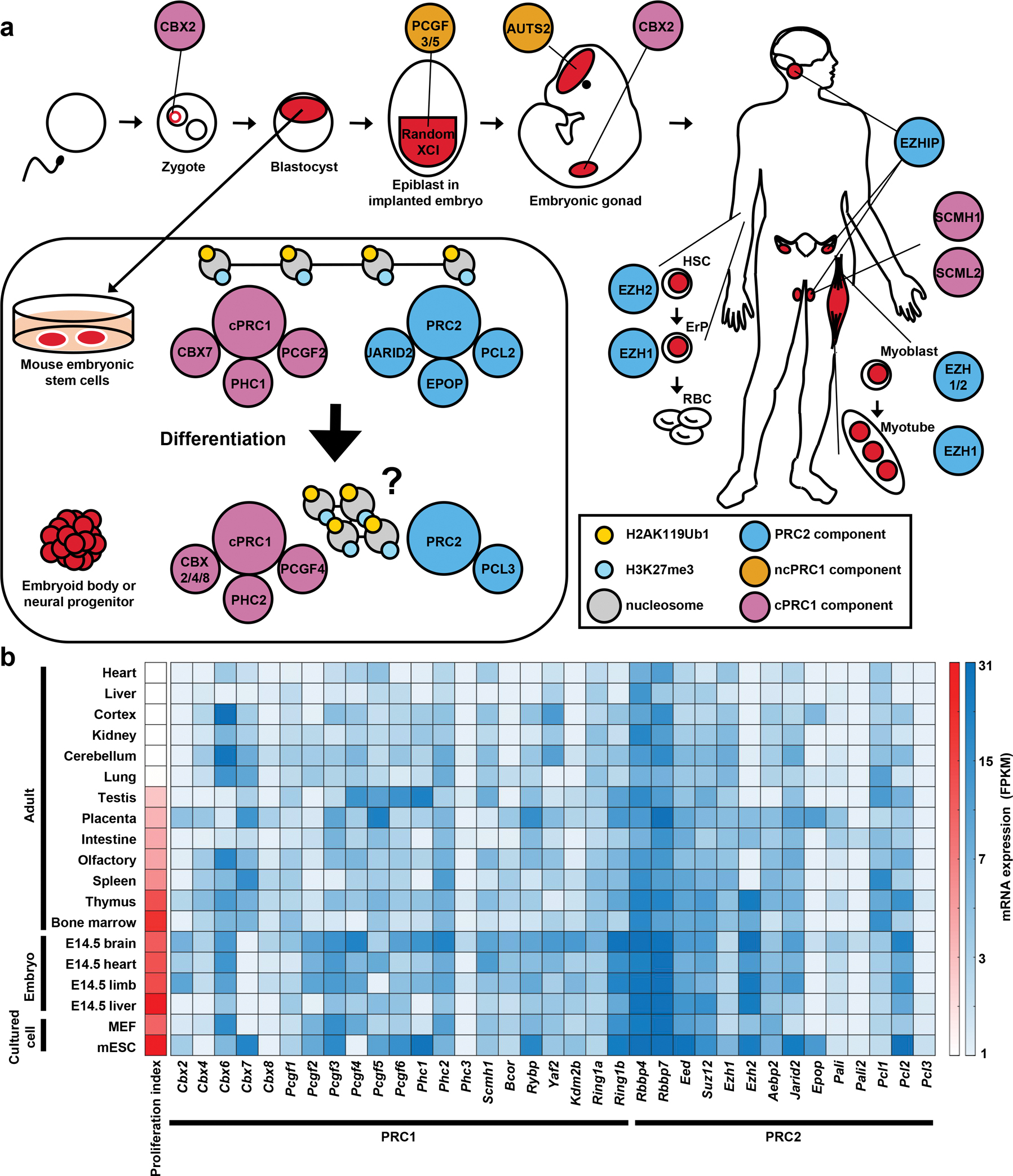
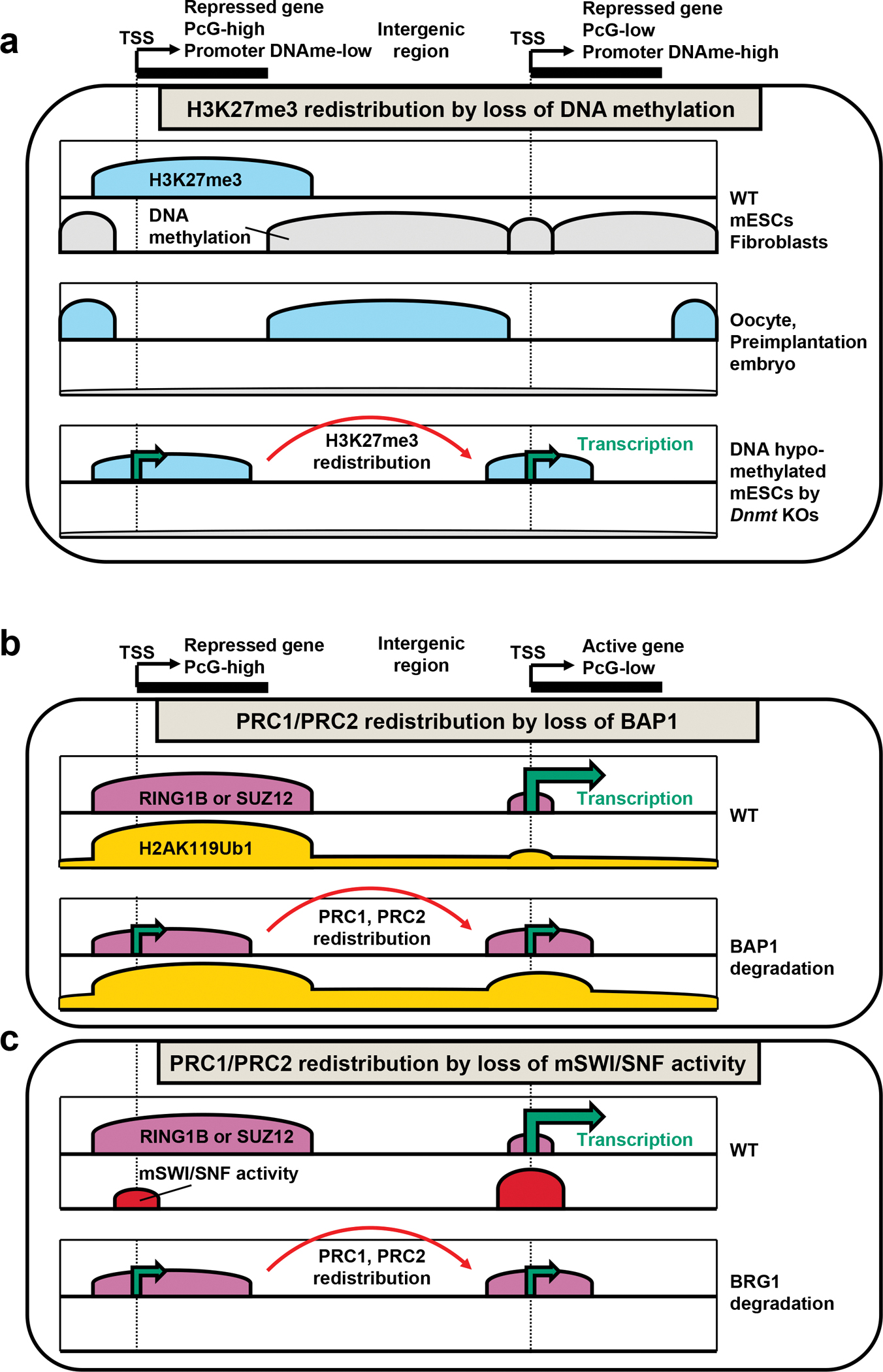
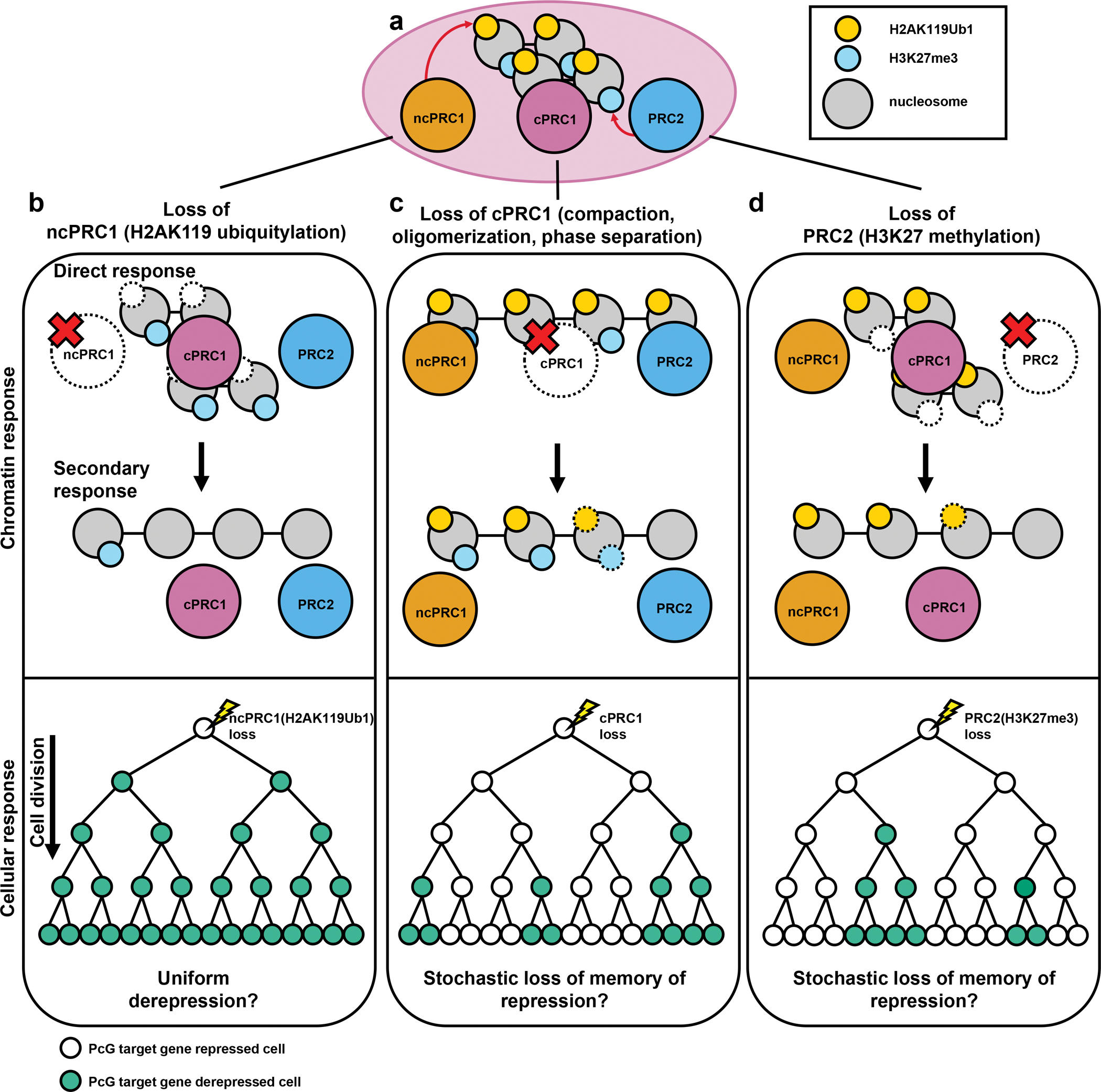
Polycomb group (PcG) proteins are crucial chromatin regulators that maintain repression of lineage-inappropriate genes and are therefore required for stable cell fate. Recent advances show that PcG proteins form distinct multi-protein complexes in various cellular environments, such as in early development, adult tissue maintenance and cancer. This surprising compositional diversity provides the basis for mechanistic diversity. Understanding this complexity deepens and refines the principles of PcG complex recruitment, target-gene repression and inheritance of memory. We review how the core molecular mechanism of Polycomb complexes operates in diverse developmental settings and propose that context-dependent changes in composition and mechanism are essential for proper epigenetic regulation in development.
© 2022. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Similar articles
-
Cell Fate and Developmental Regulation Dynamics by Polycomb Proteins and 3D Genome Architecture.Bioessays. 2019 Mar;41(3):e1800222. doi: 10.1002/bies.201800222. Epub 2019 Feb 22. Bioessays. 2019. PMID: 30793782 Review.
-
Nucleotide substitutions revealing specific functions of Polycomb group genes.Mol Genet Metab. 2015 Apr;114(4):547-56. doi: 10.1016/j.ymgme.2015.01.007. Epub 2015 Jan 30. Mol Genet Metab. 2015. PMID: 25669595 Review.
-
Combinatorial interactions are required for the efficient recruitment of pho repressive complex (PhoRC) to polycomb response elements.PLoS Genet. 2014 Jul 10;10(7):e1004495. doi: 10.1371/journal.pgen.1004495. eCollection 2014 Jul. PLoS Genet. 2014. PMID: 25010632 Free PMC article.
-
Polycomb complexes in normal and malignant hematopoiesis.J Cell Biol. 2019 Jan 7;218(1):55-69. doi: 10.1083/jcb.201808028. Epub 2018 Oct 19. J Cell Biol. 2019. PMID: 30341152 Free PMC article. Review.
-
Roles of Polycomb complexes in regulating gene expression and chromatin structure in plants.Plant Commun. 2021 Nov 26;3(1):100267. doi: 10.1016/j.xplc.2021.100267. eCollection 2022 Jan 10. Plant Commun. 2021. PMID: 35059633 Free PMC article. Review.
Cited by
-
Sparse CBX2 nucleates many Polycomb proteins to promote facultative heterochromatinization of Polycomb target genes.bioRxiv [Preprint]. 2024 Feb 5:2024.02.05.578969. doi: 10.1101/2024.02.05.578969. bioRxiv. 2024. PMID: 38370615 Free PMC article. Preprint.
-
H3K27 dimethylation dynamics reveal stepwise establishment of facultative heterochromatin in early mouse embryos.Nat Cell Biol. 2025 Jan;27(1):28-38. doi: 10.1038/s41556-024-01553-1. Epub 2024 Oct 31. Nat Cell Biol. 2025. PMID: 39482357
-
Epigenetic (De)regulation in Prostate Cancer.Cancer Treat Res. 2023;190:321-360. doi: 10.1007/978-3-031-45654-1_10. Cancer Treat Res. 2023. PMID: 38113006 Free PMC article.
-
Nuclear mRNA decay: regulatory networks that control gene expression.Nat Rev Genet. 2024 Oct;25(10):679-697. doi: 10.1038/s41576-024-00712-2. Epub 2024 Apr 18. Nat Rev Genet. 2024. PMID: 38637632 Free PMC article. Review.
-
New genetic and epigenetic insights into the chemokine system: the latest discoveries aiding progression toward precision medicine.Cell Mol Immunol. 2023 Jul;20(7):739-776. doi: 10.1038/s41423-023-01032-x. Epub 2023 May 17. Cell Mol Immunol. 2023. PMID: 37198402 Free PMC article. Review.
References
-
- Lewis PH Pc: Polycomb. Drosoph Inf Serv 21, 69 (1947).
-
- Slifer EH A mutant stock of Drosophila with extra sex-combs. Journal of Experimental Zoology 90, 31–40, doi:10.1002/jez.1400900103 (1942). - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
