

Genetic Determinants for Virulence and Transmission of the Panzootic Avian Influenza Virus H5N8 Clade 2.3.4.4 in Pekin Ducks
- PMID: 35670594
- PMCID: PMC9278104
- DOI: 10.1128/jvi.00149-22
Genetic Determinants for Virulence and Transmission of the Panzootic Avian Influenza Virus H5N8 Clade 2.3.4.4 in Pekin Ducks
Abstract
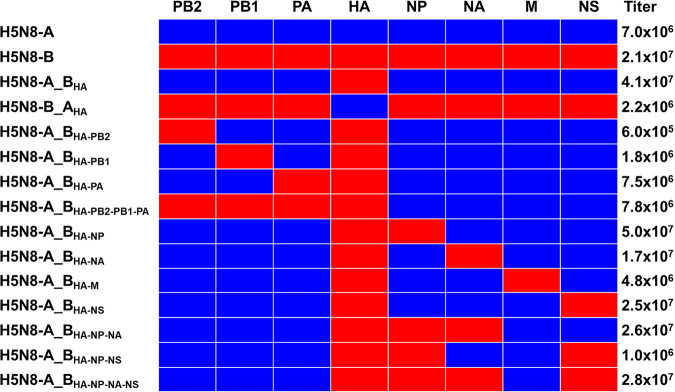
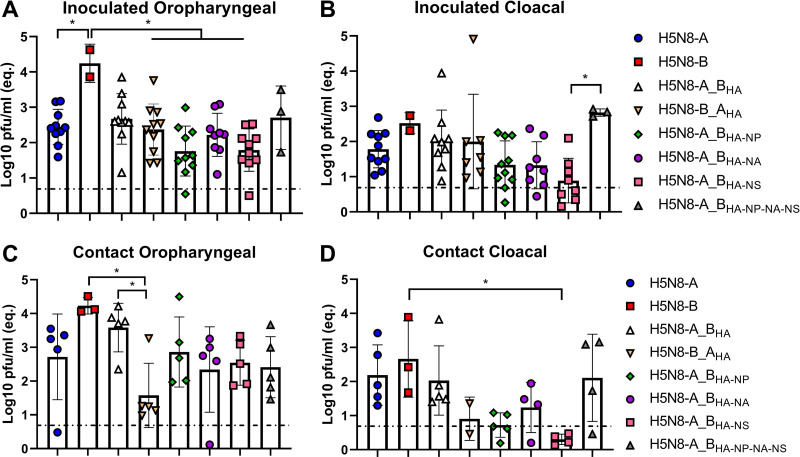
Waterfowl is the natural reservoir for avian influenza viruses (AIV), where the infection is mostly asymptomatic. In 2016, the panzootic high pathogenicity (HP) AIV H5N8 of clade 2.3.4.4B (designated H5N8-B) caused significant mortality in wild and domestic ducks, in stark contrast to the predecessor 2.3.4.4A virus from 2014 (designated H5N8-A). Here, we studied the genetic determinants for virulence and transmission of H5N8 clade 2.3.4.4 in Pekin ducks. While ducks inoculated with recombinant H5N8-A did not develop any clinical signs, H5N8-B-inoculated and cohoused ducks died after showing neurological signs. Swapping of the HA gene segments did not increase virulence of H5N8-A but abolished virulence and reduced systemic replication of H5N8-B. Only H5N8-A carrying H5N8-B HA, NP, and NS with or without NA exhibited high virulence in inoculated and contact ducks, similar to H5N8-B. Compared to H5N8-A, HA, NA, NS, and NP proteins of H5N8-B possess peculiar differences, which conferred increased receptor binding affinity, neuraminidase activity, efficiency to inhibit interferon-alpha induction, and replication in vitro, respectively. Taken together, this comprehensive study showed that HA is not the only virulence determinant of the panzootic H5N8-B in Pekin ducks, but NP, NS, and to a lesser extent NA were also necessary for the exhibition of high virulence in vivo. These proteins acted synergistically to increase receptor binding affinity, sialidase activity, interferon antagonism, and replication. This is the first ad-hoc study to investigate the mechanism underlying the high virulence of HPAIV in Pekin ducks. IMPORTANCE Since 2014, several waves of avian influenza virus (AIV) H5N8 of clade 2.3.4.4 occurred globally on unprecedented levels. Unlike viruses in the first wave in 2014-2015 (H5N8-A), viruses in 2015-2016 (H5N8-B) exhibited unusually high pathogenicity (HP) in wild and domestic ducks. Here, we found that the high virulence of H5N8-B in Pekin ducks could be attributed to multiple factors in combination, namely, hemagglutinin (HA), neuraminidase (NA), nucleoprotein (NP), and nonstructural protein 1 (NS1). Compared to H5N8-A, H5N8-B possesses distinct genetic and biological properties including increased HA receptor-binding affinity and neuraminidase activity. Likewise, H5N8-B NS1 and NP were more efficient to inhibit interferon induction and enhance replication in primary duck cells, respectively. These results indicate the polygenic trait of virulence of HPAIV in domestic ducks and the altered biological properties of the HPAIV H5N8 clade 2.3.4.4B. These findings may explain the unusual high mortality in Pekin ducks during the panzootic H5N8 outbreaks.
Keywords: H5N8; HPAI; ducks; hemagglutinin; highly pathogenic avian influenza virus; neuramindase; non-structural protein NS1; nucleoprotein; virulence determinants.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Preferential Selection and Contribution of Non-Structural Protein 1 (NS1) to the Efficient Transmission of Panzootic Avian Influenza H5N8 Virus Clades 2.3.4.4A and B in Chickens and Ducks.J Virol. 2021 Aug 25;95(18):e0044521. doi: 10.1128/JVI.00445-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34160261 Free PMC article.
-
Multiple Gene Segments Are Associated with Enhanced Virulence of Clade 2.3.4.4 H5N8 Highly Pathogenic Avian Influenza Virus in Mallards.J Virol. 2021 Aug 25;95(18):e0095521. doi: 10.1128/JVI.00955-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34232725 Free PMC article.
-
Evaluation of three hemagglutinin-based vaccines for the experimental control of a panzootic clade 2.3.4.4b A(H5N8) high pathogenicity avian influenza virus in mule ducks.Vaccine. 2023 Jan 4;41(1):145-158. doi: 10.1016/j.vaccine.2022.11.012. Epub 2022 Nov 18. Vaccine. 2023. PMID: 36411134
-
The C-terminus of non-structural protein 1 (NS1) in H5N8 clade 2.3.4.4 avian influenza virus affects virus fitness in human cells and virulence in mice.Emerg Microbes Infect. 2021 Dec;10(1):1760-1776. doi: 10.1080/22221751.2021.1971568. Emerg Microbes Infect. 2021. PMID: 34420477 Free PMC article.
-
The genetics of highly pathogenic avian influenza viruses of subtype H5 in Germany, 2006-2020.Transbound Emerg Dis. 2021 May;68(3):1136-1150. doi: 10.1111/tbed.13843. Epub 2020 Sep 29. Transbound Emerg Dis. 2021. PMID: 32964686 Review.
Cited by
-
Highly Pathogenic Avian Influenza (HPAI) H5 Clade 2.3.4.4b Virus Infection in Birds and Mammals.Animals (Basel). 2024 May 2;14(9):1372. doi: 10.3390/ani14091372. Animals (Basel). 2024. PMID: 38731377 Free PMC article. Review.
-
Virulence and transmission characteristics of clade 2.3.4.4b H5N6 subtype avian influenza viruses possessing different internal gene constellations.Virulence. 2023 Dec;14(1):2250065. doi: 10.1080/21505594.2023.2250065. Virulence. 2023. PMID: 37635408 Free PMC article.
-
Complex N-glycans are important for interspecies transmission of H7 influenza A viruses.J Virol. 2024 Apr 16;98(4):e0194123. doi: 10.1128/jvi.01941-23. Epub 2024 Mar 12. J Virol. 2024. PMID: 38470143 Free PMC article.
-
A Comparison of Host Responses to Infection with Wild-Type Avian Influenza Viruses in Chickens and Tufted Ducks.Microbiol Spectr. 2023 Aug 17;11(4):e0258622. doi: 10.1128/spectrum.02586-22. Epub 2023 Jun 26. Microbiol Spectr. 2023. PMID: 37358408 Free PMC article.
-
Conserved Expression and Functionality of Furin between Chickens and Ducks as an Activating Protease of Highly Pathogenic Avian Influenza Virus Hemagglutinins.Microbiol Spectr. 2023 Mar 14;11(2):e0460222. doi: 10.1128/spectrum.04602-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36916982 Free PMC article.
References
-
- Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith D, Rimmelzwaan GF, Olsen B, Osterhaus AD. 2005. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol 79:2814–2822. 10.1128/JVI.79.5.2814-2822.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
