

SARS-CoV-2 N Protein Antagonizes Stress Granule Assembly and IFN Production by Interacting with G3BPs to Facilitate Viral Replication
- PMID: 35652658
- PMCID: PMC9215227
- DOI: 10.1128/jvi.00412-22
SARS-CoV-2 N Protein Antagonizes Stress Granule Assembly and IFN Production by Interacting with G3BPs to Facilitate Viral Replication
Abstract
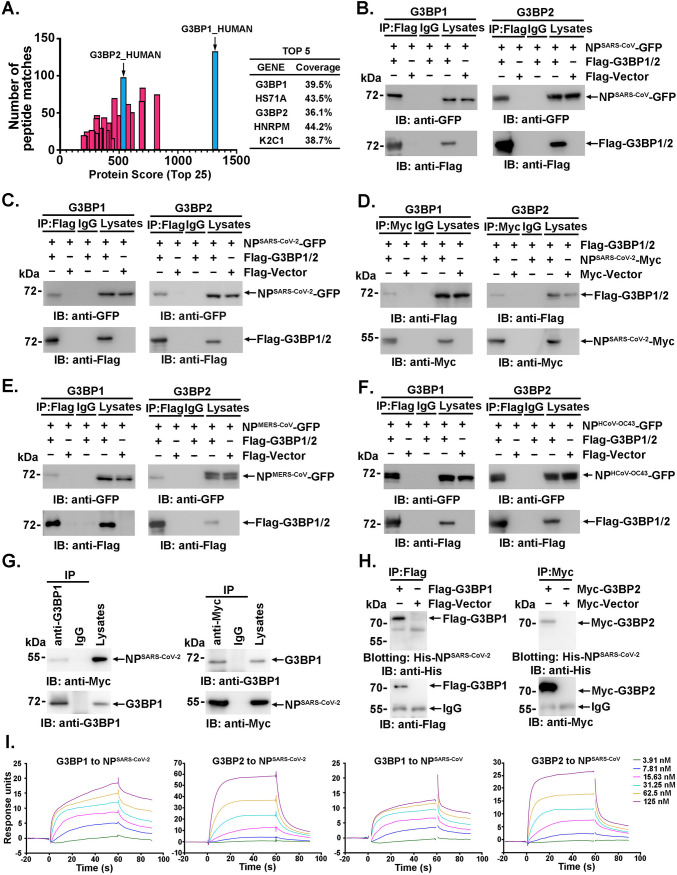
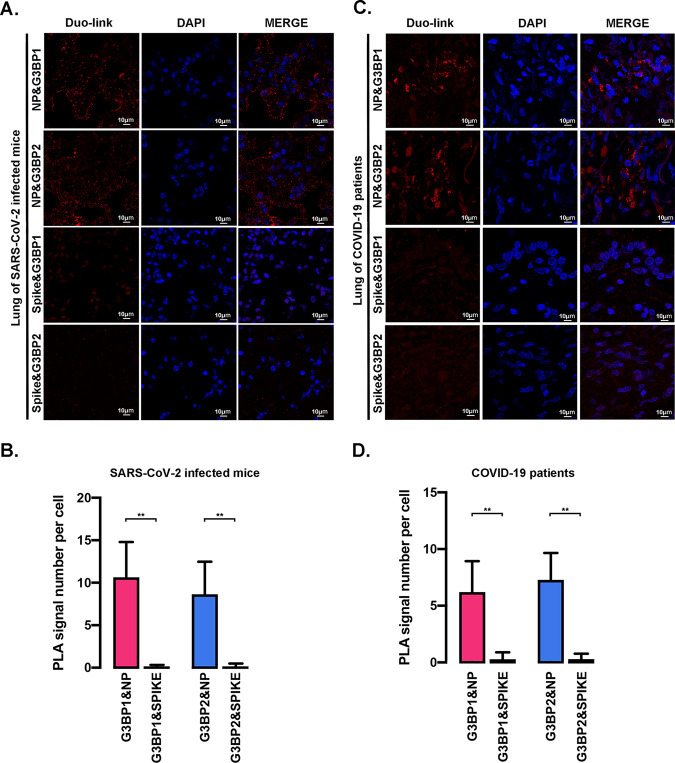
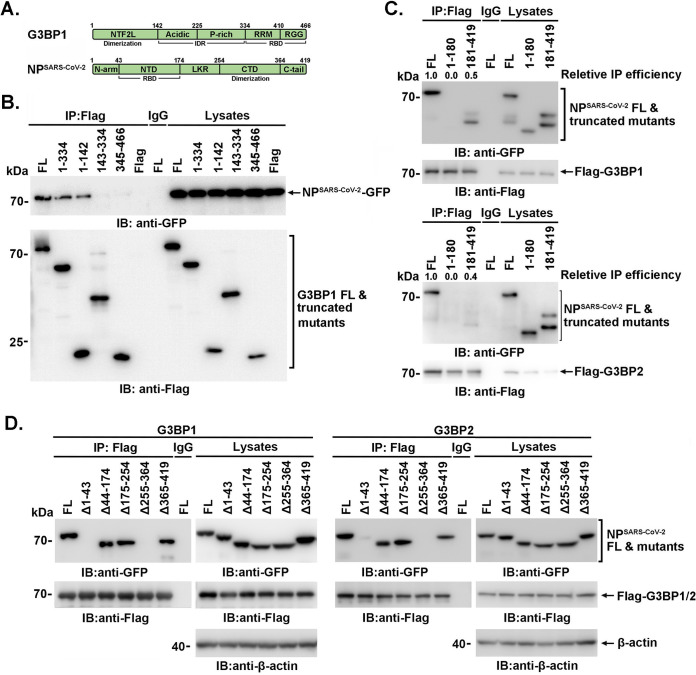
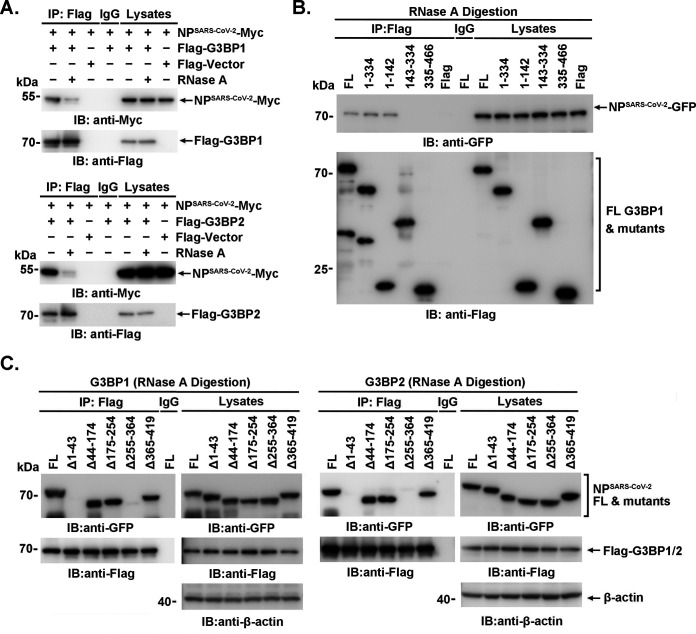
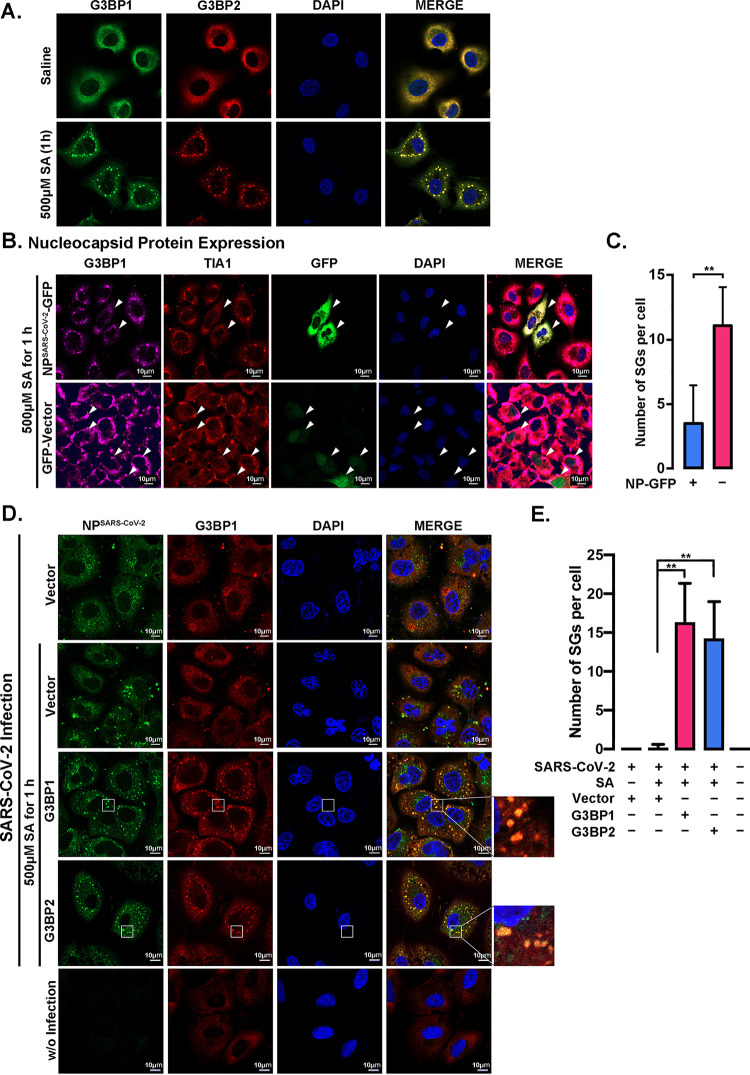
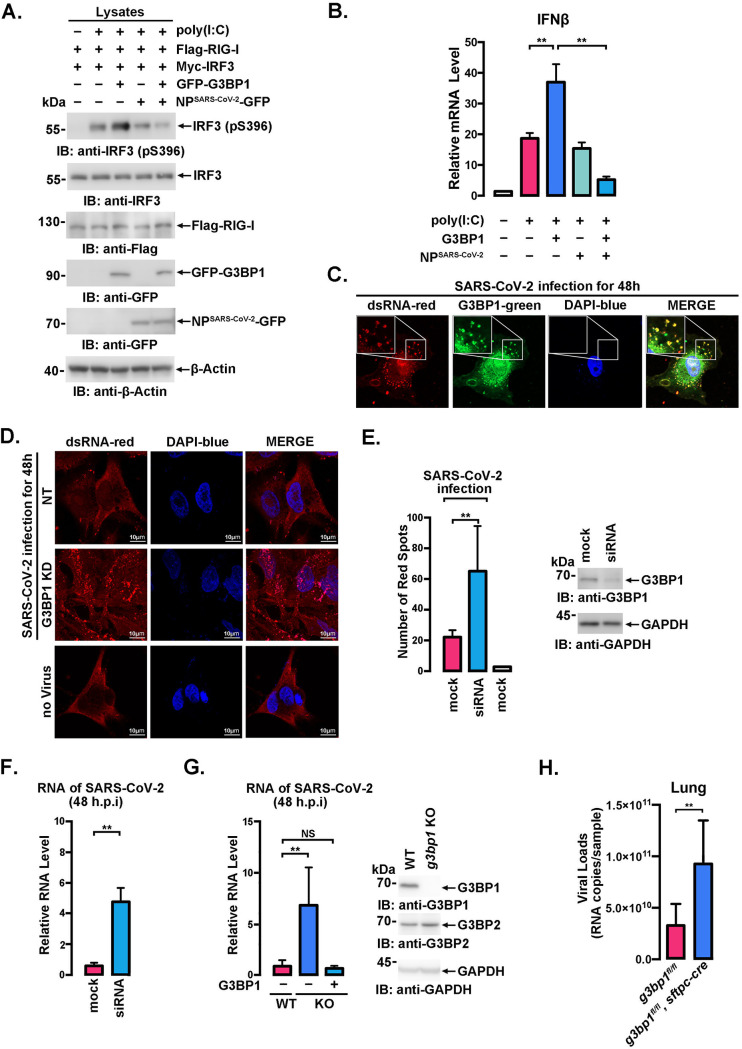
SARS-CoV-2 is the causative agent of the ongoing pandemic of coronavirus disease 2019 (COVID-19) and poses a significant threat to global health. N protein (NP), which is a major pathogenic protein among betacoronaviruses, binds to the viral RNA genome to allow viral genome packaging and viral particle release. Recent studies showed that NP antagonizes interferon (IFN) induction and mediates phase separation. Using live SARS-CoV-2 viruses, this study provides solid evidence showing that SARS-CoV-2 NP associates with G3BP1 and G3BP2 in vitro and in vivo. NPSARS-CoV-2 could efficiently suppress G3BP-mediated SG formation and potentiate viral infection by overcoming G3BP1-mediated antiviral innate immunity. G3BP1 conditional knockout mice (g3bp1fl/fL, Sftpc-Cre) exhibit significantly higher lung viral loads after SARS-CoV-2 infection than wild-type mice. Our findings contribute to the growing body of knowledge regarding the pathogenicity of NPSARS-CoV-2 and provide insight into new therapeutics targeting NPSARS-CoV-2. IMPORTANCE In this study, by in vitro assay and live SARS-CoV-2 virus infection, we provide solid evidence that the SARS-CoV-2 NP associates with G3BP1 and G3BP2 in vitro and in vivo. NPSARS-CoV-2 could efficiently suppress G3BP-mediated SG formation and potentiate viral infection by overcoming antiviral innate immunity mediated by G3BP1 in A549 cell lines and G3BP1 conditional knockout mice (g3bp1-cKO) mice, which provide in-depth evidence showing the mechanism underlying NP-related SARS-CoV-2 pathogenesis through G3BPs.
Keywords: G3BP1; SARS-CoV-2; nucleocapsid protein; stress granule.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Genomics-guided targeting of stress granule proteins G3BP1/2 to inhibit SARS-CoV-2 propagation.Int J Biol Macromol. 2021 Nov 1;190:636-648. doi: 10.1016/j.ijbiomac.2021.09.018. Epub 2021 Sep 10. Int J Biol Macromol. 2021. PMID: 34517025 Free PMC article.
-
SARS-CoV-2 nucleocapsid protein interacts with immunoregulators and stress granules and phase separates to form liquid droplets.FEBS Lett. 2021 Dec;595(23):2872-2896. doi: 10.1002/1873-3468.14229. Epub 2021 Nov 22. FEBS Lett. 2021. PMID: 34780058 Free PMC article.
-
SARS-CoV-2 Nucleocapsid Protein Antagonizes GADD34-Mediated Innate Immune Pathway through Atypical Foci.Molecules. 2024 Oct 10;29(20):4792. doi: 10.3390/molecules29204792. Molecules. 2024. PMID: 39459161 Free PMC article.
-
A closer look at mammalian antiviral condensates.Biochem Soc Trans. 2024 Jun 26;52(3):1393-1404. doi: 10.1042/BST20231296. Biochem Soc Trans. 2024. PMID: 38778761 Free PMC article. Review.
-
The roles of G3BP1 in human diseases (review).Gene. 2022 May 5;821:146294. doi: 10.1016/j.gene.2022.146294. Epub 2022 Feb 14. Gene. 2022. PMID: 35176431 Review.
Cited by
-
Role(s) of G3BPs in Human Pathogenesis.J Pharmacol Exp Ther. 2023 Oct;387(1):100-110. doi: 10.1124/jpet.122.001538. Epub 2023 Jul 19. J Pharmacol Exp Ther. 2023. PMID: 37468286 Free PMC article. Review.
-
Defining Distinct RNA-Protein Interactomes of SARS-CoV-2 Genomic and Subgenomic RNAs.J Proteome Res. 2024 Jan 5;23(1):149-160. doi: 10.1021/acs.jproteome.3c00506. Epub 2023 Dec 3. J Proteome Res. 2024. PMID: 38043095 Free PMC article.
-
Cellular Stress Responses against Coronavirus Infection: A Means of the Innate Antiviral Defense.J Microbiol Biotechnol. 2024 Jan 28;34(1):1-9. doi: 10.4014/jmb.2307.07038. Epub 2023 Sep 8. J Microbiol Biotechnol. 2024. PMID: 37674398 Free PMC article. Review.
-
SARS-CoV-2 nucleocapsid protein promotes self-deacetylation by inducing HDAC6 to facilitate viral replication.Virol J. 2024 Aug 12;21(1):186. doi: 10.1186/s12985-024-02460-5. Virol J. 2024. PMID: 39135075 Free PMC article.
-
The RNA Interference Effector Protein Argonaute 2 Functions as a Restriction Factor Against SARS-CoV-2.J Mol Biol. 2023 Aug 15;435(16):168170. doi: 10.1016/j.jmb.2023.168170. Epub 2023 Jun 3. J Mol Biol. 2023. PMID: 37271493 Free PMC article.
References
-
- Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, Si HR, Zhu Y, Li B, Huang CL, Chen HD, Chen J, Luo Y, Guo H, Jiang RD, Liu MQ, Chen Y, Shen XR, Wang X, Zheng XS, Zhao K, Chen QJ, Deng F, Liu LL, Yan B, Zhan FX, Wang YY, Xiao GF, Shi ZL. 2020. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579:270–273. 10.1038/s41586-020-2012-7. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
