

T cells discriminate between groups C1 and C2 HLA-C
- PMID: 35587797
- PMCID: PMC9177145
- DOI: 10.7554/eLife.75670
T cells discriminate between groups C1 and C2 HLA-C
Abstract
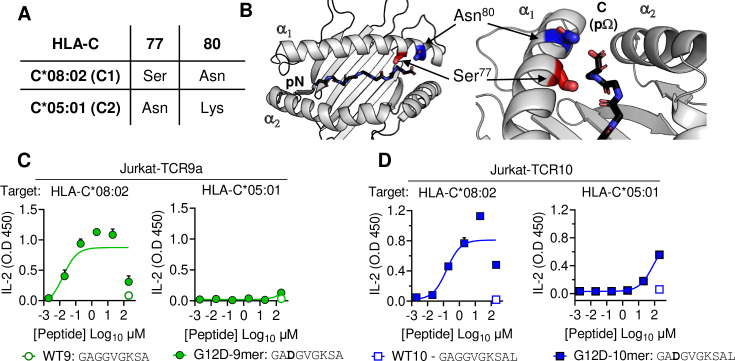
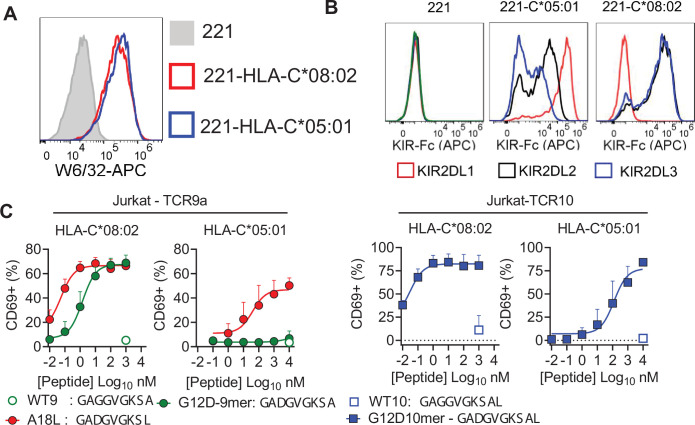
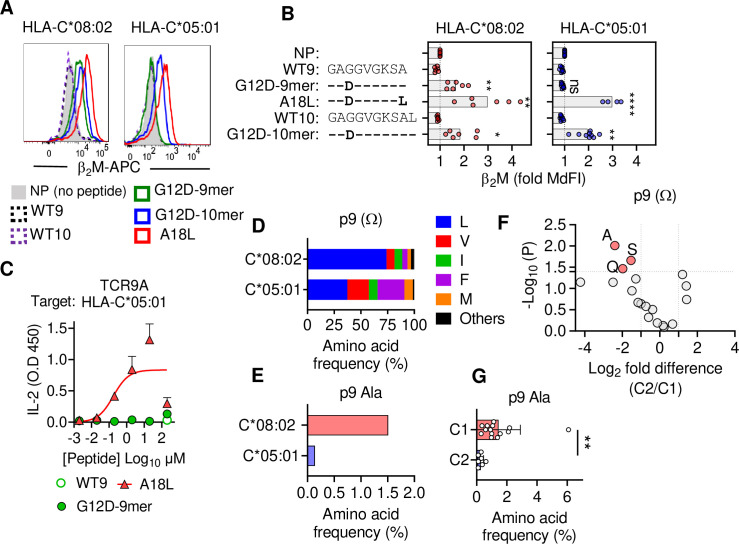
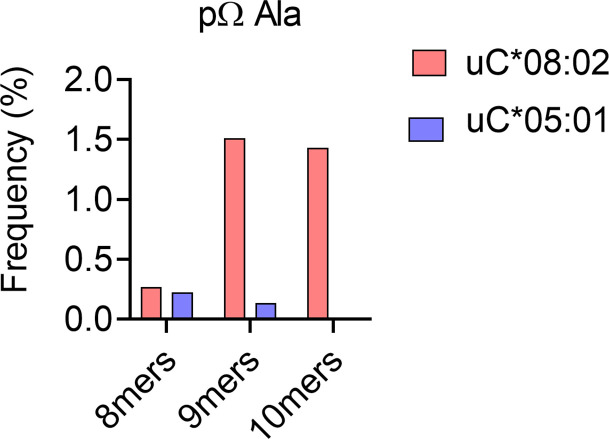
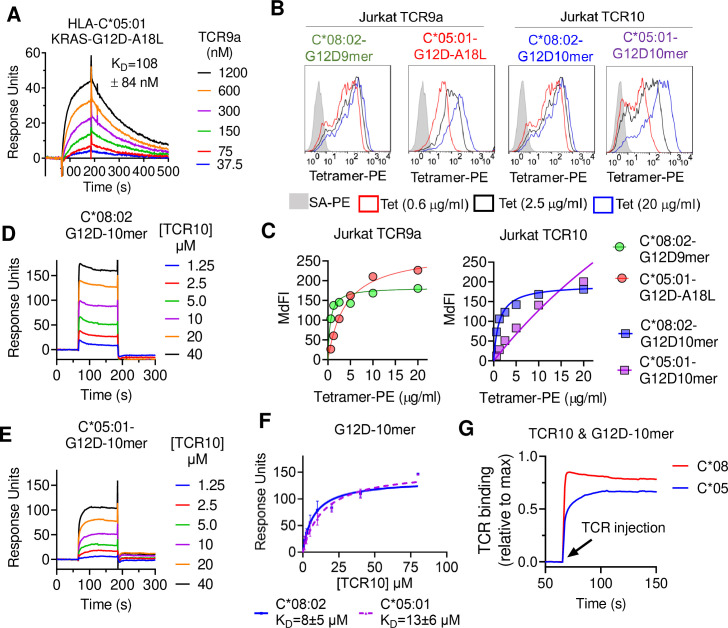
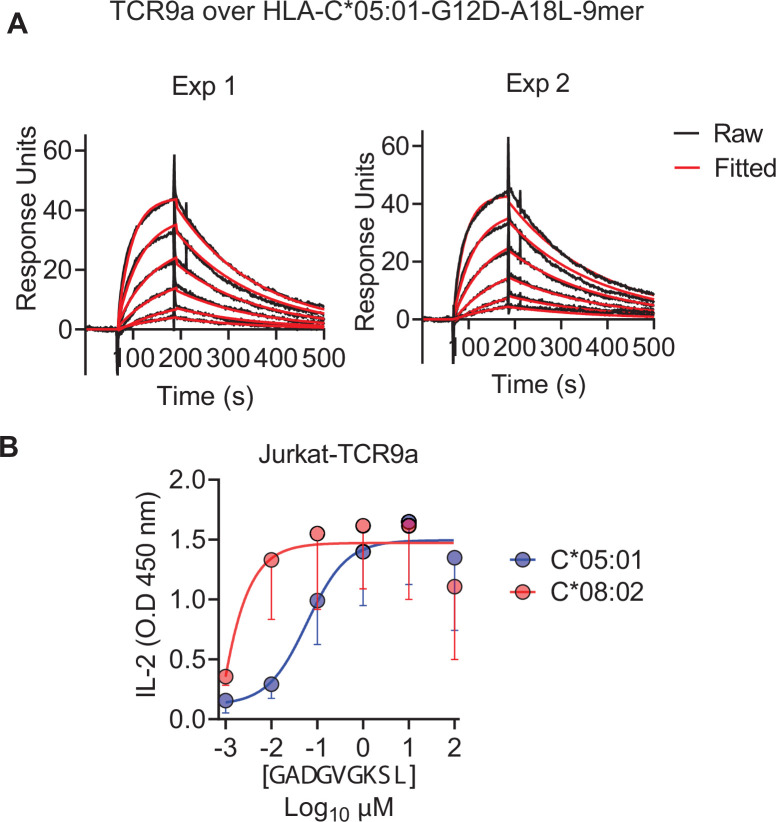
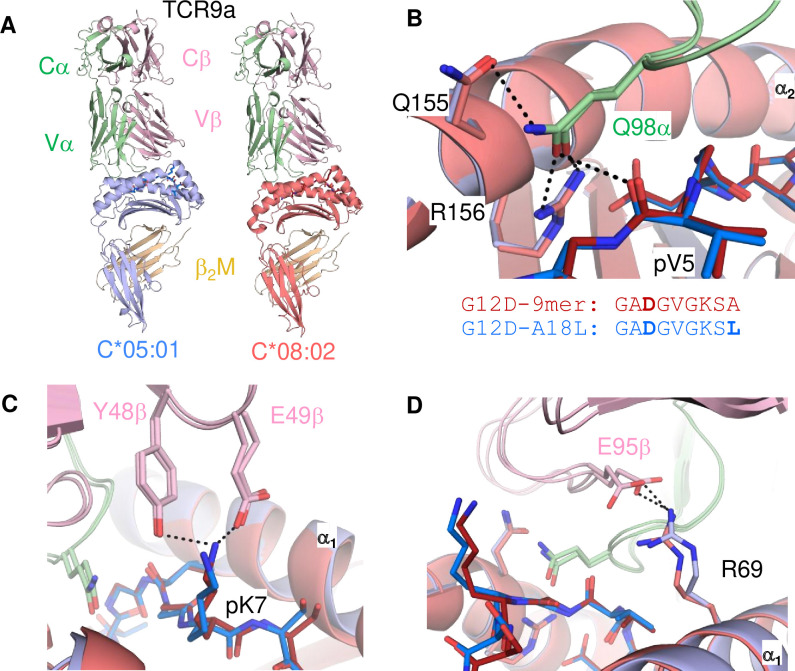
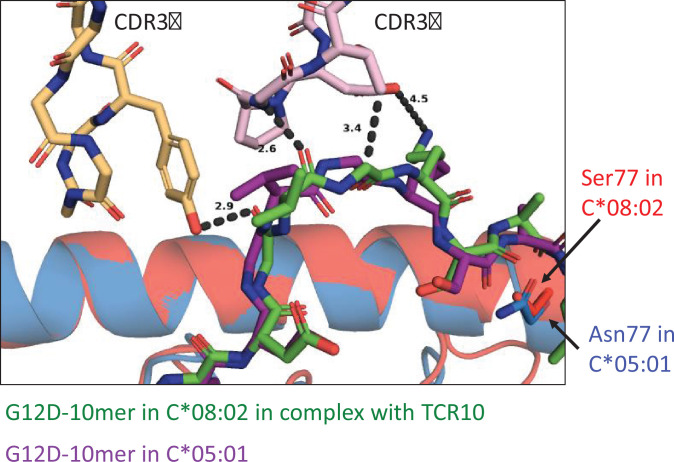
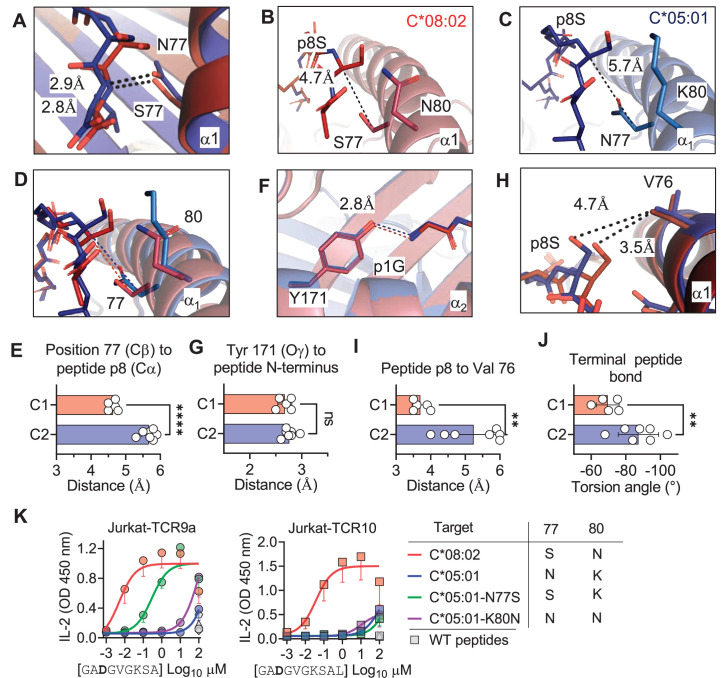
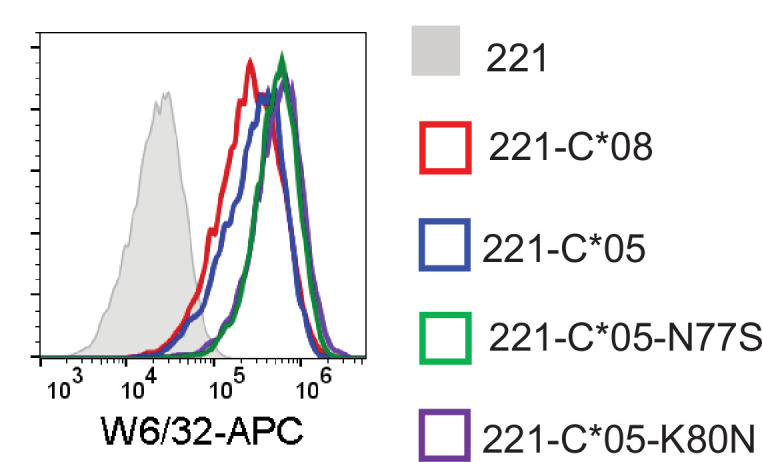
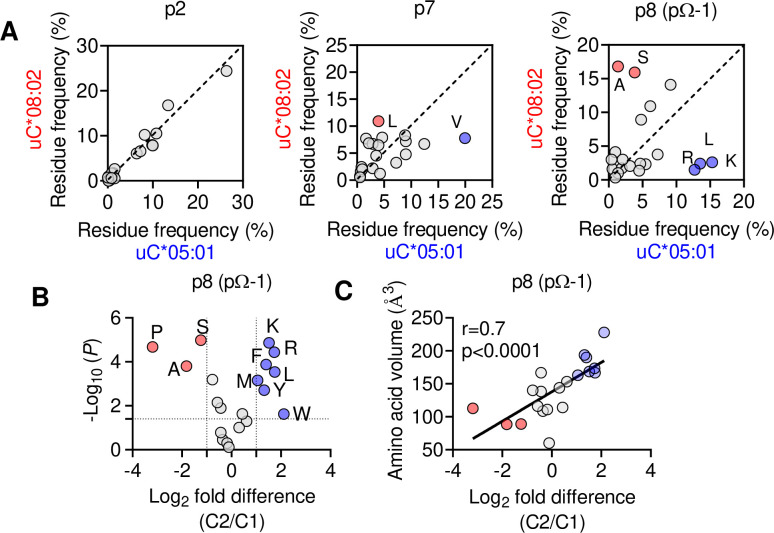
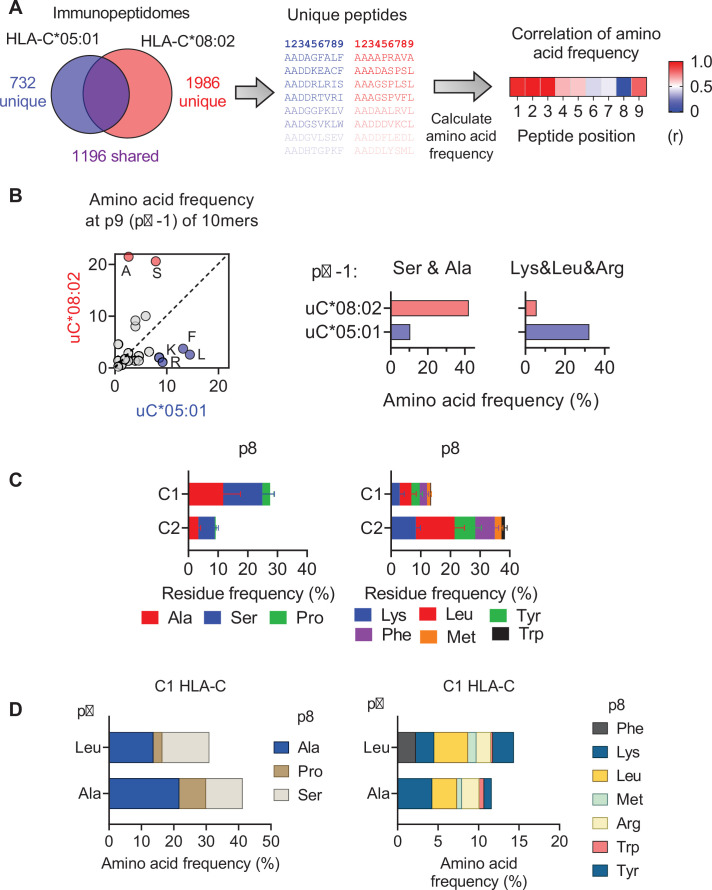
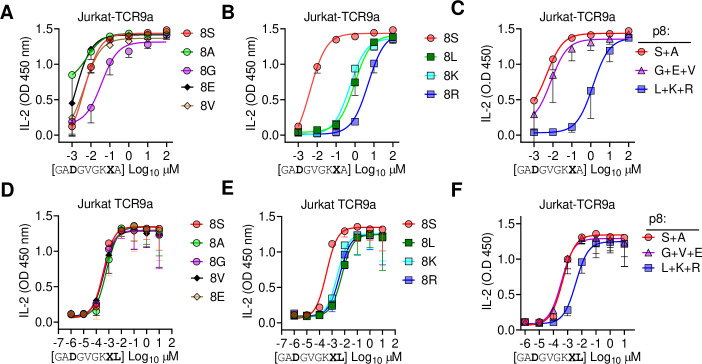
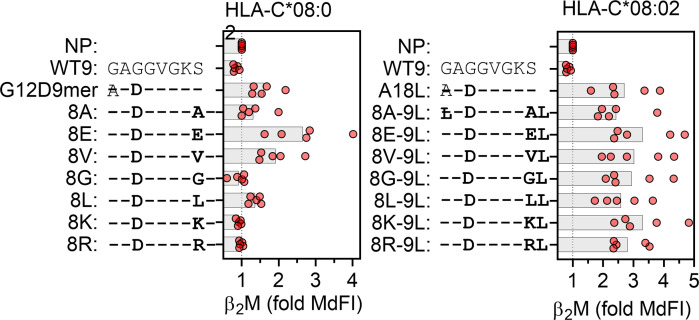
Dimorphic amino acids at positions 77 and 80 delineate HLA-C allotypes into two groups, C1 and C2, which associate with disease through interactions with C1 and C2-specific natural killer cell receptors. How the C1/C2 dimorphism affects T cell recognition is unknown. Using HLA-C allotypes that differ only by the C1/C2-defining residues, we found that KRAS-G12D neoantigen-specific T cell receptors (TCRs) discriminated between C1 and C2 presenting the same KRAS-G12D peptides. Structural and functional experiments, and immunopeptidomics analysis revealed that Ser77 in C1 and Asn77 in C2 influence amino acid preference near the peptide C-terminus (pΩ), including the pΩ-1 position, in which C1 favors small and C2 prefers large residues. This resulted in weaker TCR affinity for KRAS-G12D-bound C2-HLA-C despite conserved TCR contacts. Thus, the C1/C2 dimorphism on its own impacts peptide presentation and HLA-C-restricted T cell responses, with implications in disease, including adoptive T cell therapy targeting KRAS-G12D-induced cancers.
Keywords: C1/C2; HLA-C; KRAS-G12D; TCR; human; immunology; inflammation; molecular biophysics; neoantigen; structural biology.
Conflict of interest statement
MS, ZS, JL, PB, EL, PS No competing interests declared
Figures
Similar articles
-
Structural basis for T cell recognition of cancer neoantigens and implications for predicting neoepitope immunogenicity.Front Immunol. 2023 Nov 17;14:1303304. doi: 10.3389/fimmu.2023.1303304. eCollection 2023. Front Immunol. 2023. PMID: 38045695 Free PMC article. Review.
-
High-affinity oligoclonal TCRs define effective adoptive T cell therapy targeting mutant KRAS-G12D.Proc Natl Acad Sci U S A. 2020 Jun 9;117(23):12826-12835. doi: 10.1073/pnas.1921964117. Epub 2020 May 27. Proc Natl Acad Sci U S A. 2020. PMID: 32461371 Free PMC article.
-
Identification and structural characterization of a mutant KRAS-G12V specific TCR restricted by HLA-A3.Eur J Immunol. 2024 Sep;54(9):e2451079. doi: 10.1002/eji.202451079. Epub 2024 Jul 18. Eur J Immunol. 2024. PMID: 39030753
-
Identification of T-cell Receptors Targeting KRAS-Mutated Human Tumors.Cancer Immunol Res. 2016 Mar;4(3):204-14. doi: 10.1158/2326-6066.CIR-15-0188. Epub 2015 Dec 23. Cancer Immunol Res. 2016. PMID: 26701267 Free PMC article.
-
KIR specificity and avidity of standard and unusual C1, C2, Bw4, Bw6 and A3/11 amino acid motifs at entire HLA:KIR interface between NK and target cells, the functional and evolutionary classification of HLA class I molecules.Int J Immunogenet. 2019 Aug;46(4):217-231. doi: 10.1111/iji.12433. Epub 2019 Jun 18. Int J Immunogenet. 2019. PMID: 31210416 Review.
Cited by
-
Unveiling the Significance of HLA and KIR Diversity in Underrepresented Populations.Biomedicines. 2024 Jun 15;12(6):1333. doi: 10.3390/biomedicines12061333. Biomedicines. 2024. PMID: 38927540 Free PMC article. Review.
-
Structural basis for T cell recognition of cancer neoantigens and implications for predicting neoepitope immunogenicity.Front Immunol. 2023 Nov 17;14:1303304. doi: 10.3389/fimmu.2023.1303304. eCollection 2023. Front Immunol. 2023. PMID: 38045695 Free PMC article. Review.
-
The Molecular Mechanisms of HLA-G Regulatory Function on Immune Cells during Early Pregnancy.Biomolecules. 2023 Aug 3;13(8):1213. doi: 10.3390/biom13081213. Biomolecules. 2023. PMID: 37627278 Free PMC article. Review.
-
T Cell Recognition of Tumor Neoantigens and Insights Into T Cell Immunotherapy.Front Immunol. 2022 Feb 10;13:833017. doi: 10.3389/fimmu.2022.833017. eCollection 2022. Front Immunol. 2022. PMID: 35222422 Free PMC article. Review.
-
HLA-C Peptide Repertoires as Predictors of Clinical Response during Early SARS-CoV-2 Infection.Life (Basel). 2024 Sep 19;14(9):1181. doi: 10.3390/life14091181. Life (Basel). 2024. PMID: 39337964 Free PMC article.
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallographica. Section D, Biological Crystallography. 2002;58:1948–1954. doi: 10.1107/s0907444902016657. - DOI - PubMed
-
- Addo MM, Altfeld M, Rosenberg ES, Eldridge RL, Philips MN, Habeeb K, Khatri A, Brander C, Robbins GK, Mazzara GP, Goulder PJ, Walker BD, HIV Controller Study Collaboration The HIV-1 regulatory proteins Tat and Rev are frequently targeted by cytotoxic T lymphocytes derived from HIV-1-infected individuals. PNAS. 2001;98:1781–1786. doi: 10.1073/pnas.98.4.1781. - DOI - PMC - PubMed
-
- Apps R, Qi Y, Carlson JM, Chen H, Gao X, Thomas R, Yuki Y, Del Prete GQ, Goulder P, Brumme ZL, Brumme CJ, John M, Mallal S, Nelson G, Bosch R, Heckerman D, Stein JL, Soderberg KA, Moody MA, Denny TN, Zeng X, Fang J, Moffett A, Lifson JD, Goedert JJ, Buchbinder S, Kirk GD, Fellay J, McLaren P, Deeks SG, Pereyra F, Walker B, Michael NL, Weintrob A, Wolinsky S, Liao W, Carrington M. Influence of HLA-C expression level on HIV control. Science (New York, N.Y.) 2013;340:87–91. doi: 10.1126/science.1232685. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
