

Refining colorectal cancer classification and clinical stratification through a single-cell atlas
- PMID: 35538548
- PMCID: PMC9092724
- DOI: 10.1186/s13059-022-02677-z
Refining colorectal cancer classification and clinical stratification through a single-cell atlas
Erratum in
-
Correction: Refining colorectal cancer classification and clinical stratification through a single-cell atlas.Genome Biol. 2022 Jul 13;23(1):156. doi: 10.1186/s13059-022-02724-9. Genome Biol. 2022. PMID: 35831907 Free PMC article. No abstract available.
Abstract
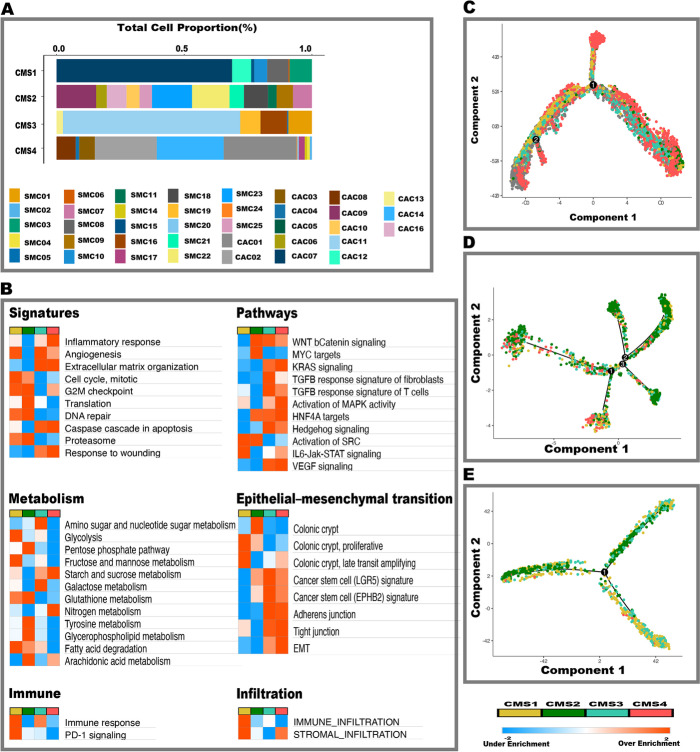
Background: Colorectal cancer (CRC) consensus molecular subtypes (CMS) have different immunological, stromal cell, and clinicopathological characteristics. Single-cell characterization of CMS subtype tumor microenvironments is required to elucidate mechanisms of tumor and stroma cell contributions to pathogenesis which may advance subtype-specific therapeutic development. We interrogate racially diverse human CRC samples and analyze multiple independent external cohorts for a total of 487,829 single cells enabling high-resolution depiction of the cellular diversity and heterogeneity within the tumor and microenvironmental cells.
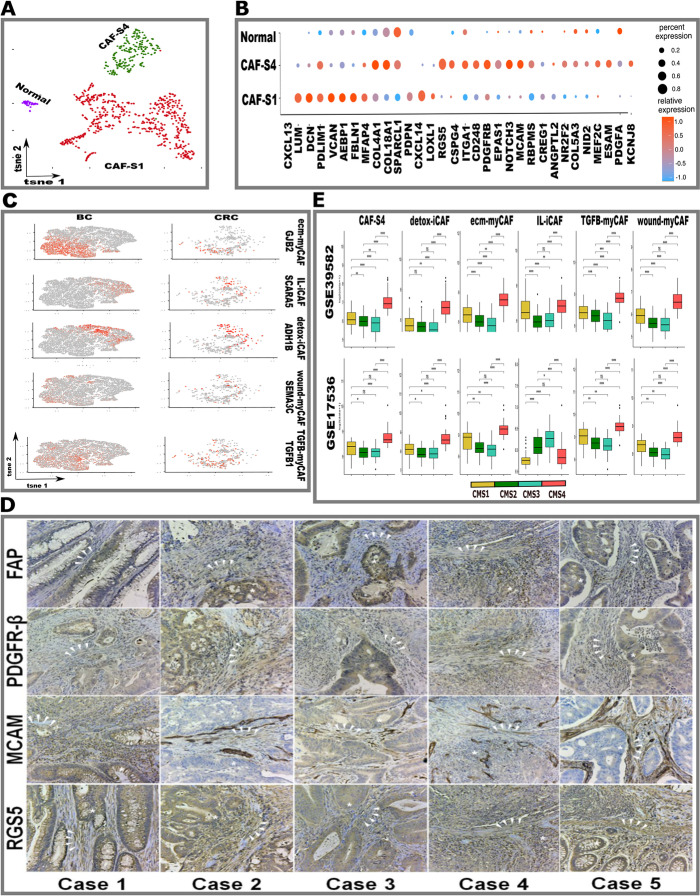
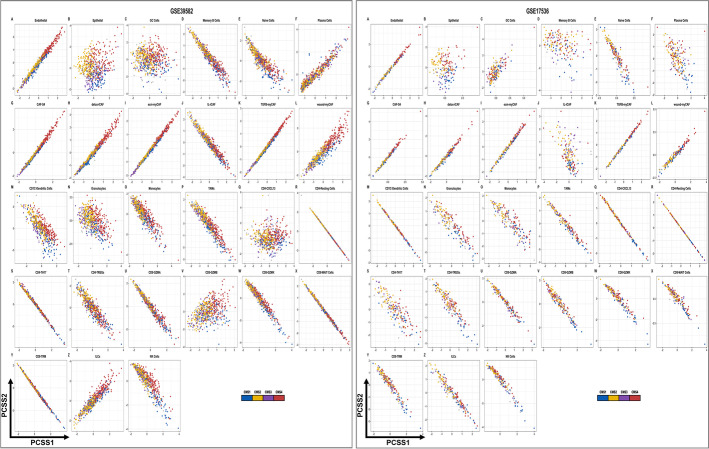
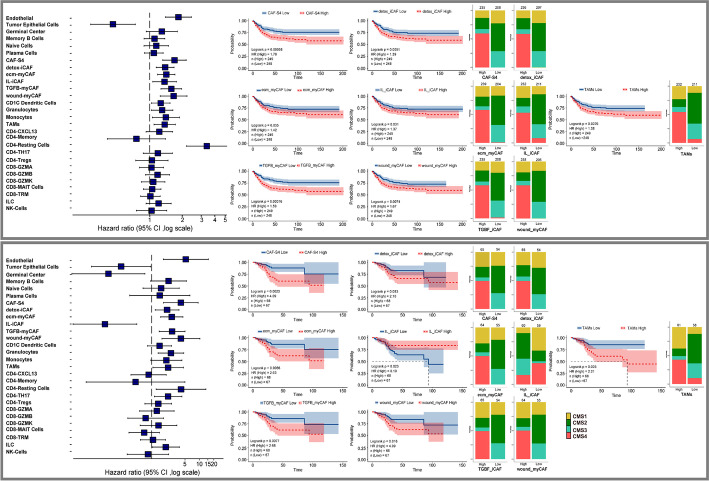
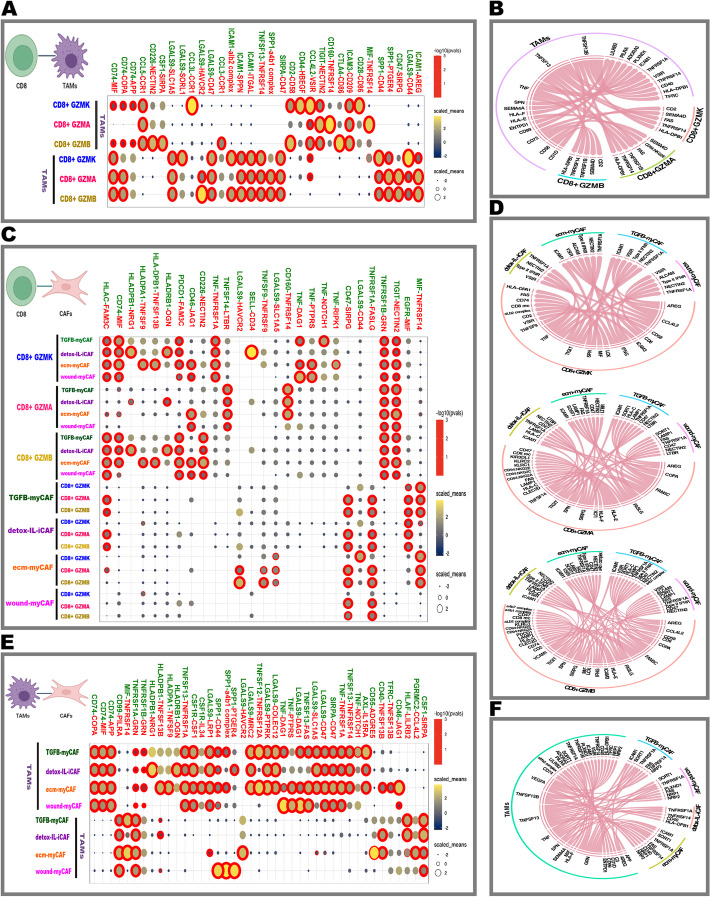
Results: Tumor cells recapitulate individual CMS subgroups yet exhibit significant intratumoral CMS heterogeneity. Both CMS1 microsatellite instability (MSI-H) CRCs and microsatellite stable (MSS) CRC demonstrate similar pathway activations at the tumor epithelial level. However, CD8+ cytotoxic T cell phenotype infiltration in MSI-H CRCs may explain why these tumors respond to immune checkpoint inhibitors. Cellular transcriptomic profiles in CRC exist in a tumor immune stromal continuum in contrast to discrete subtypes proposed by studies utilizing bulk transcriptomics. We note a dichotomy in tumor microenvironments across CMS subgroups exists by which patients with high cancer-associated fibroblasts (CAFs) and C1Q+TAM content exhibit poor outcomes, providing a higher level of personalization and precision than would distinct subtypes. Additionally, we discover CAF subtypes known to be associated with immunotherapy resistance.
Conclusions: Distinct CAFs and C1Q+ TAMs are sufficient to explain CMS predictive ability and a simpler signature based on these cellular phenotypes could stratify CRC patient prognosis with greater precision. Therapeutically targeting specific CAF subtypes and C1Q + TAMs may promote immunotherapy responses in CRC patients.
Keywords: CMS classification; Cancer-associated fibroblast; Colorectal cancer; Immunotherapy; Single-cell analysis; Stromal signatures.
© 2022. The Author(s).
Conflict of interest statement
Ashiq Masood and Jeffrey A. Borgia received research funding from Tempus lab. Anjuraj Sadanandam receives research funding from Bristol-Myers Squibb; Merck KGaA, Pierre Fabre. Furthermore, Anjuraj Sadanandam. holds patent PCT/IB2013/060416, “Colorectal cancer classification with differential prognosis and personalized therapeutic responses,” and patent number 2011213.2 “Prognostic and Treatment Response Predictive Method.”
Figures



Similar articles
-
Relative contribution of clinicopathological variables, genomic markers, transcriptomic subtyping and microenvironment features for outcome prediction in stage II/III colorectal cancer.Ann Oncol. 2019 Oct 1;30(10):1622-1629. doi: 10.1093/annonc/mdz287. Ann Oncol. 2019. PMID: 31504112 Free PMC article.
-
Crosstalk Between the MSI Status and Tumor Microenvironment in Colorectal Cancer.Front Immunol. 2020 Aug 12;11:2039. doi: 10.3389/fimmu.2020.02039. eCollection 2020. Front Immunol. 2020. PMID: 32903444 Free PMC article.
-
Genomic and transcriptomic characterization of heterogeneous immune subgroups of microsatellite instability-high colorectal cancers.J Immunother Cancer. 2021 Dec;9(12):e003414. doi: 10.1136/jitc-2021-003414. J Immunother Cancer. 2021. PMID: 34903553 Free PMC article.
-
The Evolving Role of Consensus Molecular Subtypes: a Step Beyond Inpatient Selection for Treatment of Colorectal Cancer.Curr Treat Options Oncol. 2021 Nov 6;22(12):113. doi: 10.1007/s11864-021-00913-5. Curr Treat Options Oncol. 2021. PMID: 34741675 Review.
-
Relationships Between Immune Landscapes, Genetic Subtypes and Responses to Immunotherapy in Colorectal Cancer.Front Immunol. 2020 Mar 6;11:369. doi: 10.3389/fimmu.2020.00369. eCollection 2020. Front Immunol. 2020. PMID: 32210966 Free PMC article. Review.
Cited by
-
NLRP12/C1qA positive feedback in tumor-associated macrophages regulates immunosuppression through LILRB4/NF-κB pathway in lung adenocarcinoma.Cancer Immunol Immunother. 2024 Nov 11;74(1):16. doi: 10.1007/s00262-024-03880-6. Cancer Immunol Immunother. 2024. PMID: 39527158 Free PMC article.
-
Molecular portraits of colorectal cancer morphological regions.Elife. 2023 Nov 13;12:RP86655. doi: 10.7554/eLife.86655. Elife. 2023. PMID: 37956043 Free PMC article.
-
Identification of an Amino Acid Metabolism Reprogramming Signature for Predicting Prognosis, Immunotherapy Efficacy, and Drug Candidates in Colon Cancer.Appl Biochem Biotechnol. 2024 Sep 2. doi: 10.1007/s12010-024-05049-4. Online ahead of print. Appl Biochem Biotechnol. 2024. PMID: 39222169
-
DrugMAP: molecular atlas and pharma-information of all drugs.Nucleic Acids Res. 2023 Jan 6;51(D1):D1288-D1299. doi: 10.1093/nar/gkac813. Nucleic Acids Res. 2023. PMID: 36243961 Free PMC article.
-
scRank infers drug-responsive cell types from untreated scRNA-seq data using a target-perturbed gene regulatory network.Cell Rep Med. 2024 Jun 18;5(6):101568. doi: 10.1016/j.xcrm.2024.101568. Epub 2024 May 15. Cell Rep Med. 2024. PMID: 38754419 Free PMC article.
References
-
- Molinari C, Marisi G, Passardi A, Matteucci L, De Maio G, Ulivi P. Heterogeneity in colorectal cancer: a challenge for personalized medicine? Int J Mol Sci. 2018;19 Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6321493/. Cited 2020 Dec 29. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
