

Sophisticated expression responses of ZNT1 and MT in response to changes in the expression of ZIPs
- PMID: 35513474
- PMCID: PMC9072671
- DOI: 10.1038/s41598-022-10925-2
Sophisticated expression responses of ZNT1 and MT in response to changes in the expression of ZIPs
Abstract
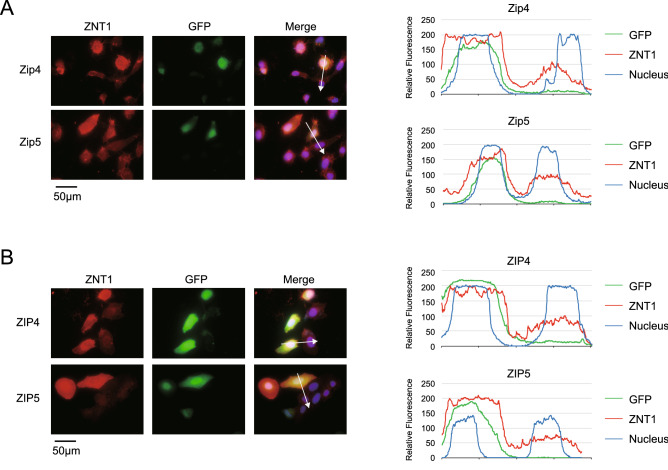
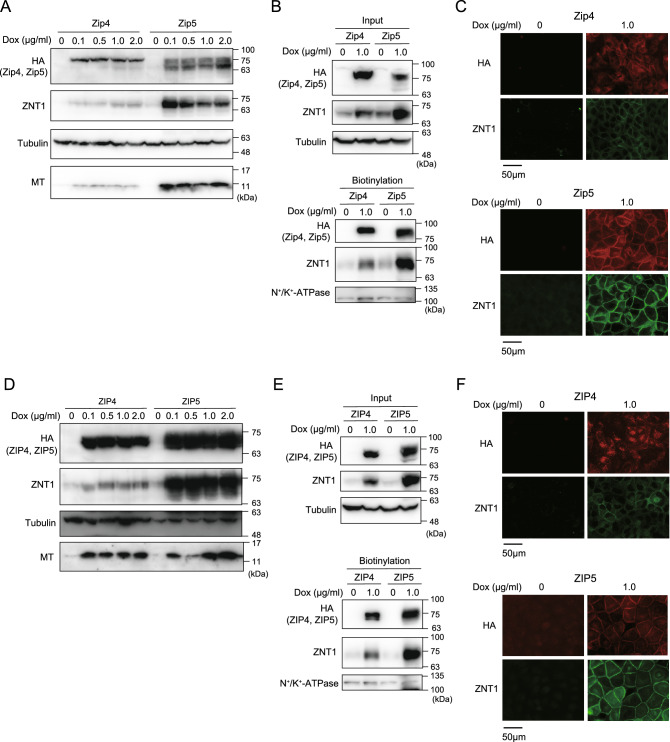
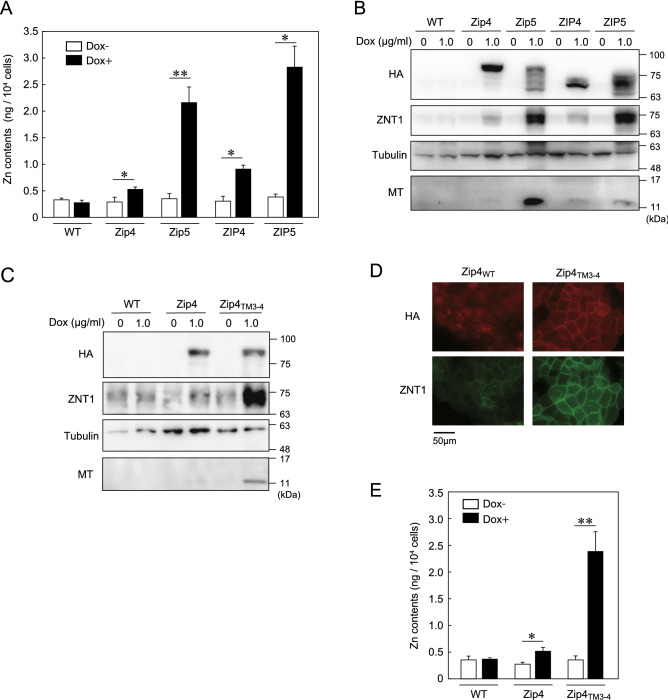
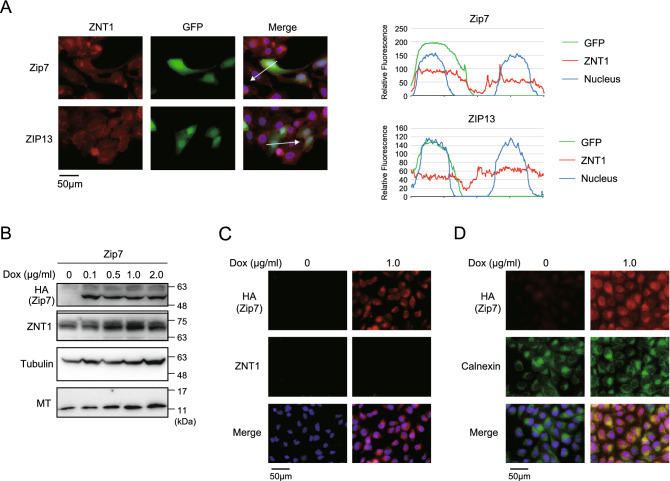
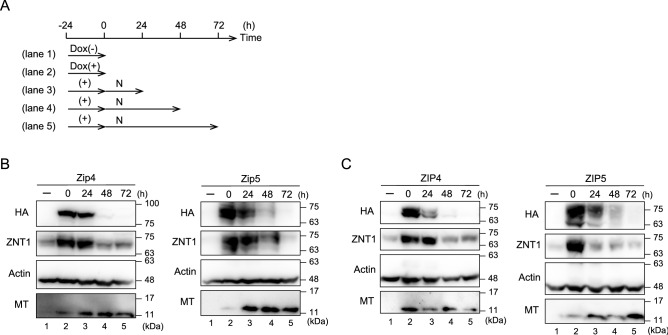
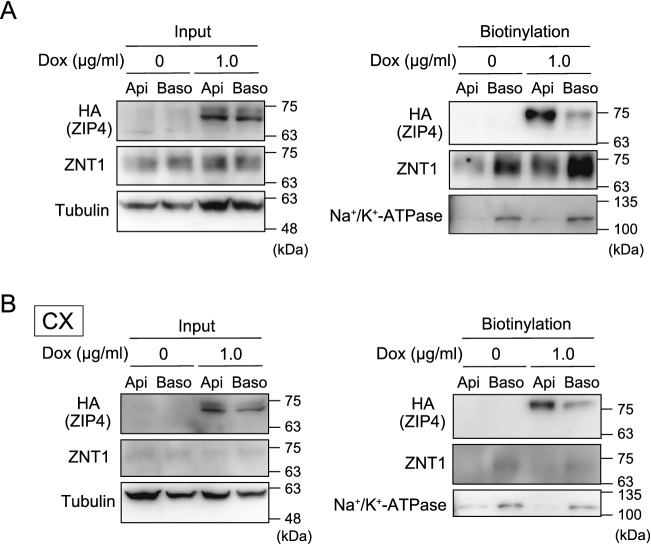
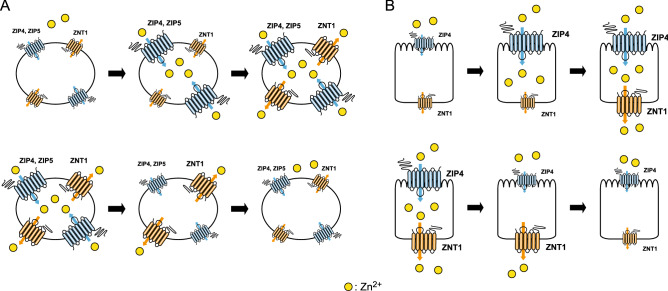
The zinc homeostatic proteins Zn transporter 1 (ZNT1) and metallothionein (MT) function in dampening increases in cytosolic zinc concentrations. Conversely, the expression of ZNT1 and MT is expected to be suppressed during decreases in cytosolic zinc concentrations. Thus, ZNT1/MT homeostatic responses are considered to be essential for maintaining cellular zinc homeostasis because cellular zinc concentrations are readily altered by changes in the expression of several Zrt-/Irt-like proteins (ZIPs) under both physiological and pathological conditions. However, this notion remains to be tested experimentally. Here, we investigated the aforementioned homeostatic process by analyzing ZNT1 and MT protein expression in response to ZIP expression. Overexpression of cell-surface-localized ZIPs, such as ZIP4 and ZIP5, increased the cellular zinc content, which caused an increase in the expression of cell-surface ZNT1 and cytosolic MT in the absence of zinc supplementation in the culture medium. By contrast, elimination of the overexpressed ZIP4 and ZIP5 resulted in decreased expression of ZNT1 but not MT, which suggests that differential regulation of ZNT1 and MT expression at the protein level underlies the homeostatic responses necessary for zinc metabolism under certain conditions. Moreover, increased expression of apically localized ZIP4 facilitated basolateral ZNT1 expression in polarized cells, which indicates that such a coordinated expression mechanism is crucial for vectorial transcellular transport. Our results provide novel insights into the physiological maintenance of cellular zinc homeostasis in response to alterations in cytosolic zinc concentrations caused by changes in the expression of ZIPs.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Simple in vitro method to evaluate ZIP zinc transport ability through zinc transporter 1 and metallothionein expression measurements.Methods Enzymol. 2023;687:207-239. doi: 10.1016/bs.mie.2023.04.017. Epub 2023 May 18. Methods Enzymol. 2023. PMID: 37666633
-
Cooperative functions of ZnT1, metallothionein and ZnT4 in the cytoplasm are required for full activation of TNAP in the early secretory pathway.PLoS One. 2013 Oct 18;8(10):e77445. doi: 10.1371/journal.pone.0077445. eCollection 2013. PLoS One. 2013. PMID: 24204829 Free PMC article.
-
Increased expression of zinc transporter ZIP4, ZIP11, ZnT1, and ZnT6 predicts poor prognosis in pancreatic cancer.J Trace Elem Med Biol. 2021 May;65:126734. doi: 10.1016/j.jtemb.2021.126734. Epub 2021 Feb 17. J Trace Elem Med Biol. 2021. PMID: 33631610
-
Gastrointestinal factors influencing zinc absorption and homeostasis.Int J Vitam Nutr Res. 2010 Oct;80(4-5):243-8. doi: 10.1024/0300-9831/a000030. Int J Vitam Nutr Res. 2010. PMID: 21462106 Free PMC article. Review.
-
Mammalian zinc transporters: nutritional and physiologic regulation.Annu Rev Nutr. 2009;29:153-76. doi: 10.1146/annurev-nutr-033009-083312. Annu Rev Nutr. 2009. PMID: 19400752 Review.
Cited by
-
Intracellular zinc signaling influences NMDA receptor function by enhancing the interaction of ZnT1 with GluN2A.Neurosci Lett. 2022 Nov 1;790:136896. doi: 10.1016/j.neulet.2022.136896. Epub 2022 Oct 3. Neurosci Lett. 2022. PMID: 36202195 Free PMC article.
-
Effects of Different Dietary Zinc (Zn) Sources on Growth Performance, Zn Metabolism, and Intestinal Health of Grass Carp.Antioxidants (Basel). 2023 Aug 23;12(9):1664. doi: 10.3390/antiox12091664. Antioxidants (Basel). 2023. PMID: 37759967 Free PMC article.
-
Modulation of Adverse Health Effects of Environmental Cadmium Exposure by Zinc and Its Transporters.Biomolecules. 2024 May 31;14(6):650. doi: 10.3390/biom14060650. Biomolecules. 2024. PMID: 38927054 Free PMC article. Review.
-
Dietary Dihydromyricetin Zinc Chelate Supplementation Improves the Intestinal Health of Magang Geese.Biol Trace Elem Res. 2024 Nov;202(11):5219-5234. doi: 10.1007/s12011-024-04065-z. Epub 2024 Jan 24. Biol Trace Elem Res. 2024. PMID: 38263355
-
Zinc's Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis.Biomolecules. 2022 Nov 11;12(11):1672. doi: 10.3390/biom12111672. Biomolecules. 2022. PMID: 36421686 Free PMC article. Review.
References
-
- Kambe T, Suzuki E, Komori T. Zinc transporters—A review and a new view from biochemistry. In: Fukada T, Kambe T, editors. Zinc Signaling. 2. Springer; 2019. pp. 23–56.
