

Abnormal triaging of misfolded proteins by adult neuronal ceroid lipofuscinosis-associated DNAJC5/CSPα mutants causes lipofuscin accumulation
- PMID: 35506243
- PMCID: PMC9809949
- DOI: 10.1080/15548627.2022.2065618
Abnormal triaging of misfolded proteins by adult neuronal ceroid lipofuscinosis-associated DNAJC5/CSPα mutants causes lipofuscin accumulation
Abstract
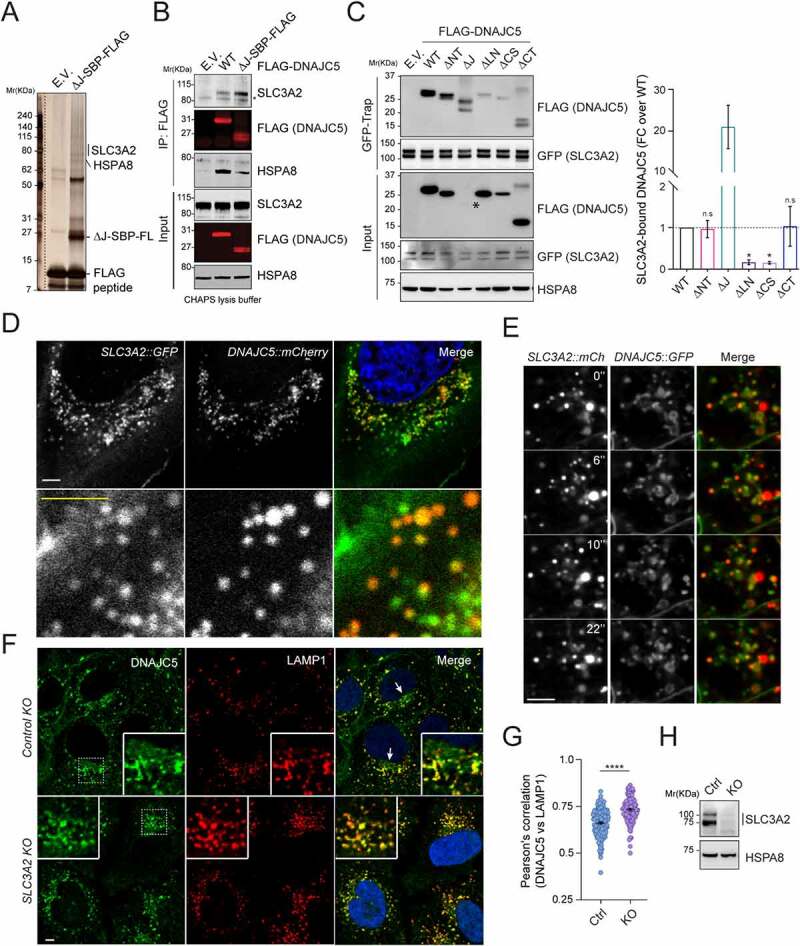
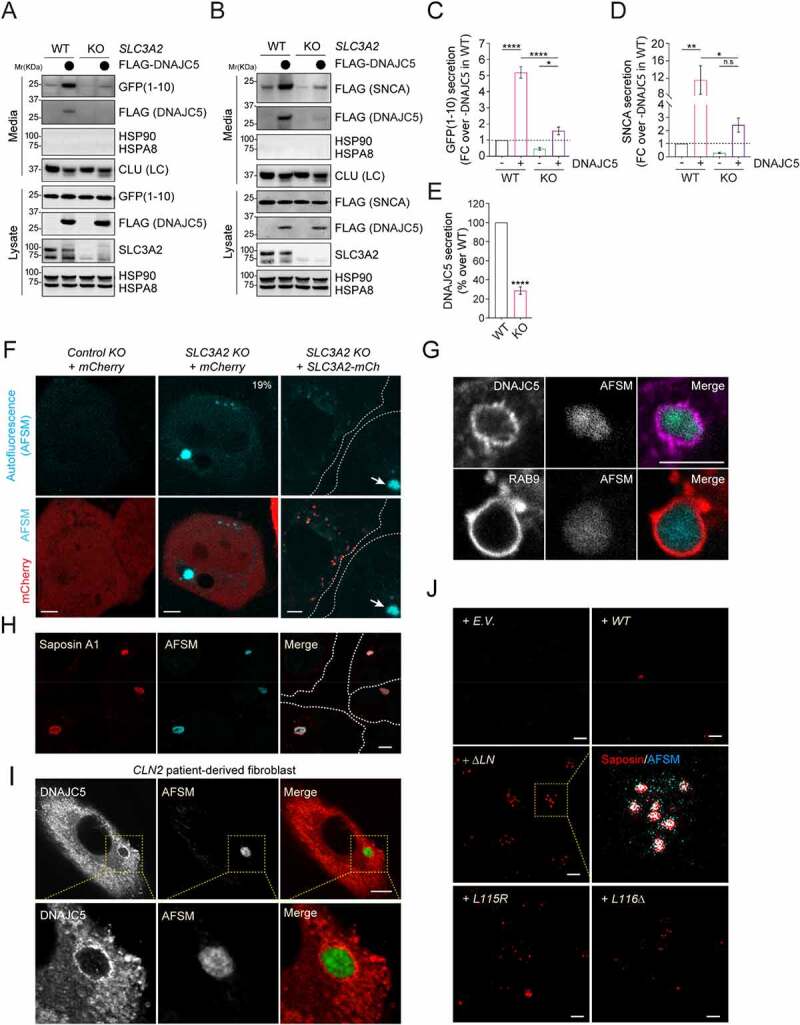
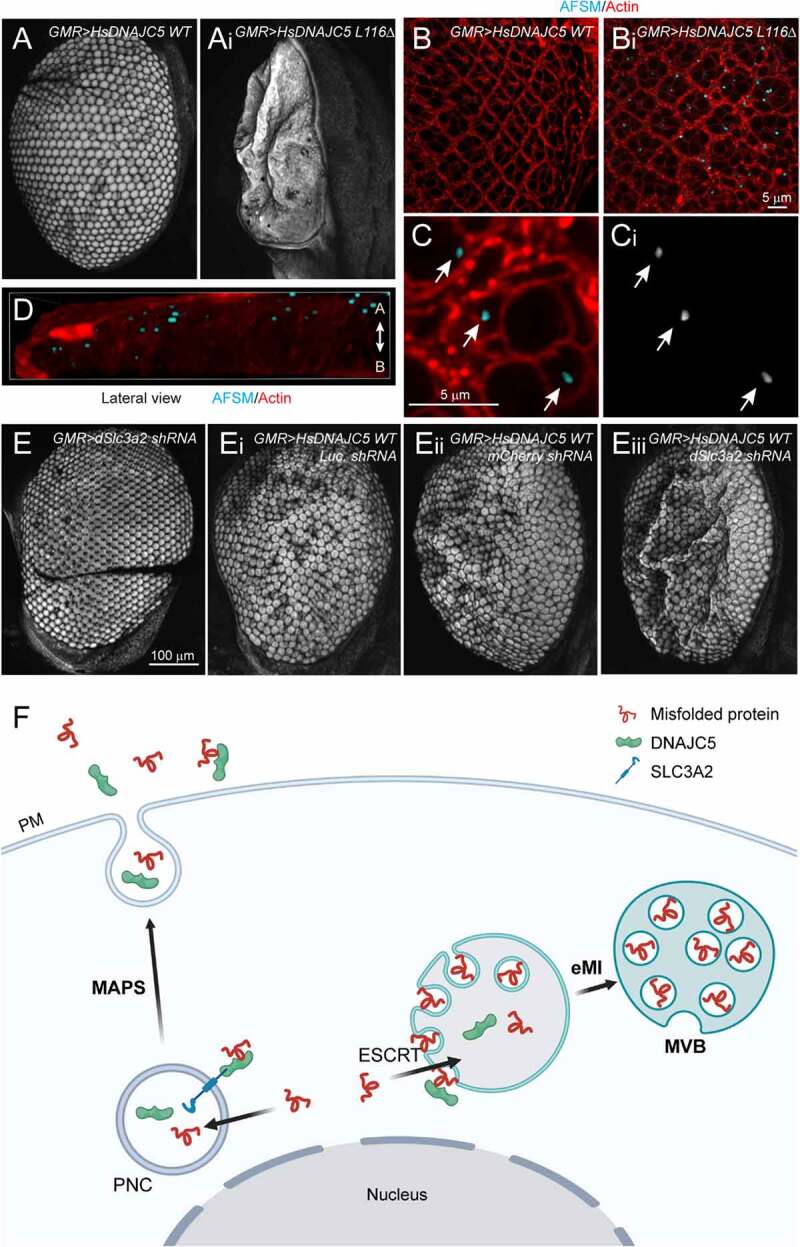
Mutations in DNAJC5/CSPα are associated with adult neuronal ceroid lipofuscinosis (ANCL), a dominant-inherited neurodegenerative disease featuring lysosome-derived autofluorescent storage materials (AFSMs) termed lipofuscin. Functionally, DNAJC5 has been implicated in chaperoning synaptic proteins and in misfolding-associated protein secretion (MAPS), but how DNAJC5 dysfunction causes lipofuscinosis and neurodegeneration is unclear. Here we report two functionally distinct but coupled chaperoning activities of DNAJC5, which jointly regulate lysosomal homeostasis: While endolysosome-associated DNAJC5 promotes ESCRT-dependent microautophagy, a fraction of perinuclear and non-lysosomal DNAJC5 mediates MAPS. Functional proteomics identifies a previously unknown DNAJC5 interactor SLC3A2/CD98hc that is essential for the perinuclear DNAJC5 localization and MAPS but dispensable for microautophagy. Importantly, uncoupling these two processes, as seen in cells lacking SLC3A2 or expressing ANCL-associated DNAJC5 mutants, generates DNAJC5-containing AFSMs resembling NCL patient-derived lipofuscin and induces neurodegeneration in a Drosophila ANCL model. These findings suggest that MAPS safeguards microautophagy to avoid DNAJC5-associated lipofuscinosis and neurodegeneration.Abbreviations: 3-MA: 3-methyladenine; ACTB: actin beta; AFSM: autofluorescent storage materials; ANCL: adult neuronal ceroid lipofuscinosis; Baf. A1: bafilomycin A1; CLN: ceroid lipofuscinosis neuronal; CLU: clusterin; CS: cysteine string domain of DNAJC5/CSPα; CUPS: compartment for unconventional protein secretion; DN: dominant negative; DNAJC5/CSPα: DnaJ heat shock protein family (Hsp40) member C5; eMI: endosomal microautophagy; ESCRT: endosomal sorting complex required for transport; GFP: green fluorescent protein; HSPA8/HSC70: heat shock protein family A (Hsp70) member 8; INCL: infant neuronal ceroid lipofuscinosis; JNCL: juvenile neuronal ceroid lipofuscinosis; KO: knockout; LAMP1: lysosomal associated membrane protein 1; LAPTM4B: lysosomal protein transmembrane 4 beta; LN: linker domain of DNAJC5/CSPα; MAPS: misfolding-associated protein secretion; mCh/Ch: mCherry; mCi/Ci: mCitrine; MTOR: mechanistic target of rapamycin kinase; NCL: neuronal ceroid lipofuscinosis; PPT1: palmitoyl-protein thioesterase 1; PQC: protein quality control; SBP: streptavidin binding protein; SGT: small glutamine-rich tetratricopeptide repeat; shRNA: short hairpin RNA; SLC3A2/CD98hc: solute carrier family 3 member 2; SNCA/α-synuclein: synuclein alpha; TMED10: transmembrane p24 trafficking protein 10; UV: ultraviolet; VPS4: vacuolar protein sorting 4 homolog; WT: wild type.
Keywords: CLN4; DNAJC5/CSPα; ESCRT; cysteine string protein α; lysobody; lysosome/endolysosome; microautophagy/eMI; misfolding-associated protein secretion/MAPS; neuronal ceroid lipofuscinosis/NCL; protein quality control/PQC; unconventional protein secretion/UPS.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
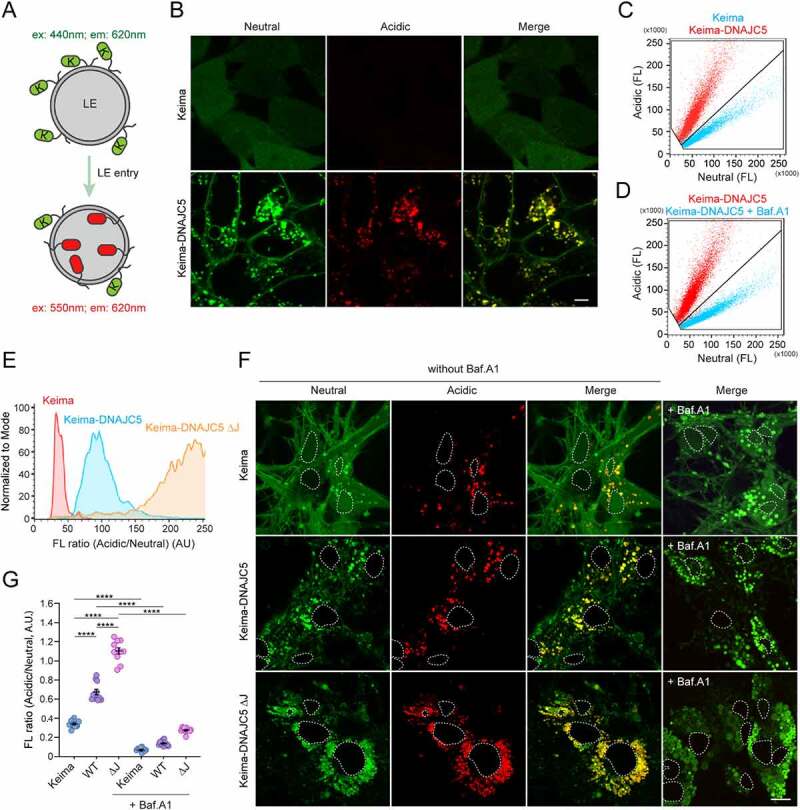
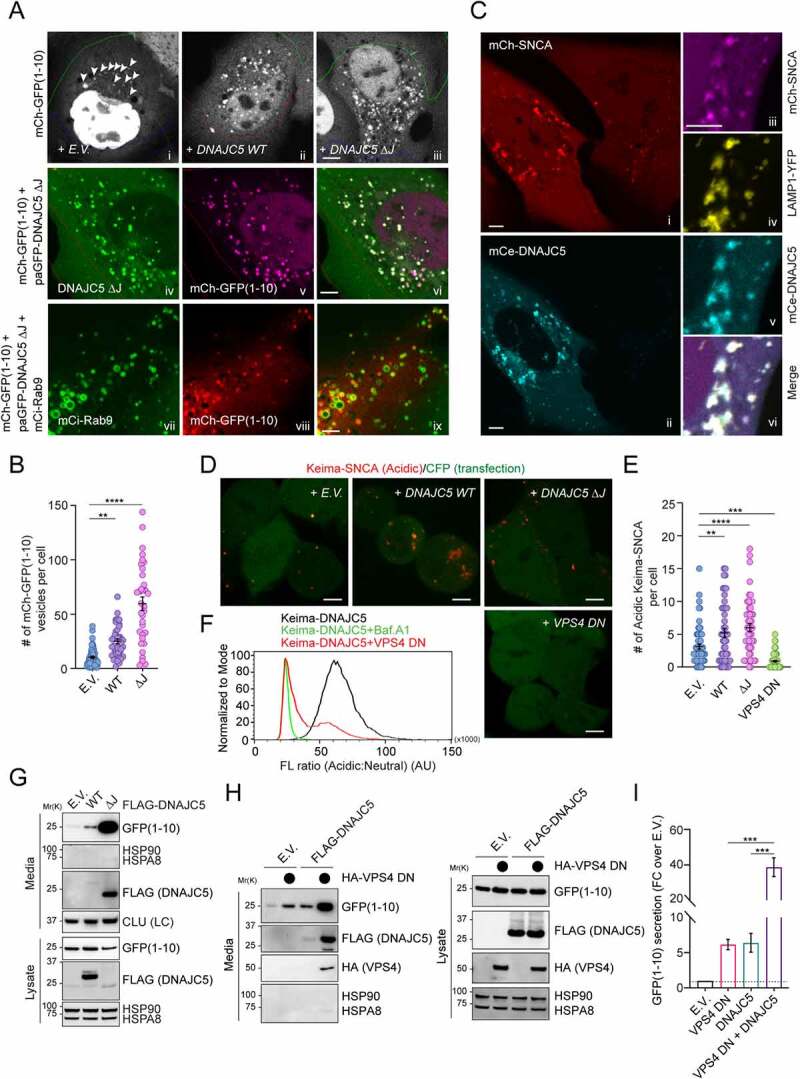
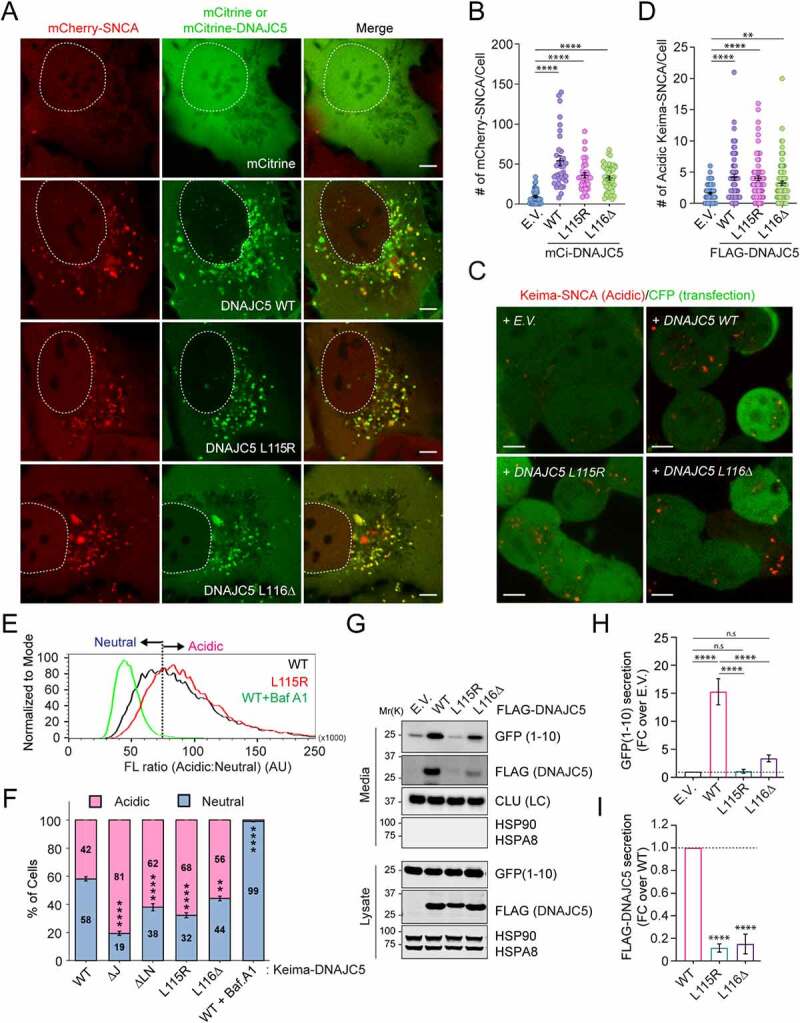
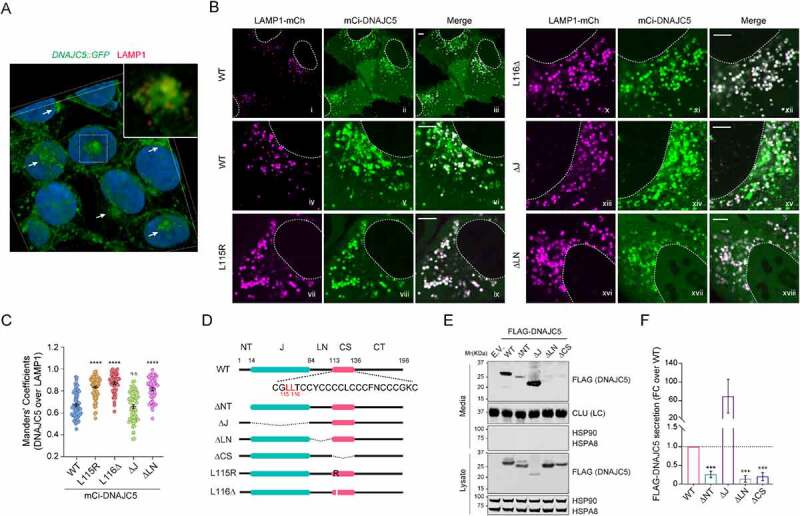
Similar articles
-
Safeguarding Lysosomal Homeostasis by DNAJC5/CSPα-Mediated Unconventional Protein Secretion and Endosomal Microautophagy.Front Cell Dev Biol. 2022 May 10;10:906453. doi: 10.3389/fcell.2022.906453. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35620055 Free PMC article. Review.
-
Neuronal ceroid lipofuscinosis with DNAJC5/CSPα mutation has PPT1 pathology and exhibit aberrant protein palmitoylation.Acta Neuropathol. 2016 Apr;131(4):621-37. doi: 10.1007/s00401-015-1512-2. Epub 2015 Dec 10. Acta Neuropathol. 2016. PMID: 26659577 Free PMC article.
-
Mutations in DNAJC5, encoding cysteine-string protein alpha, cause autosomal-dominant adult-onset neuronal ceroid lipofuscinosis.Am J Hum Genet. 2011 Aug 12;89(2):241-52. doi: 10.1016/j.ajhg.2011.07.003. Epub 2011 Aug 4. Am J Hum Genet. 2011. PMID: 21820099 Free PMC article.
-
Autosomal-dominant adult neuronal ceroid lipofuscinosis caused by duplication in DNAJC5 initially missed by Sanger and whole-exome sequencing.Eur J Hum Genet. 2020 Jun;28(6):783-789. doi: 10.1038/s41431-019-0567-2. Epub 2020 Jan 9. Eur J Hum Genet. 2020. PMID: 31919451 Free PMC article.
-
Autosomal dominant neuronal ceroid lipofuscinosis: Clinical features and molecular basis.Clin Genet. 2021 Jan;99(1):111-118. doi: 10.1111/cge.13829. Epub 2020 Aug 26. Clin Genet. 2021. PMID: 32783189 Free PMC article. Review.
Cited by
-
Converging links between adult-onset neurodegenerative Alzheimer's disease and early life neurodegenerative neuronal ceroid lipofuscinosis?Neural Regen Res. 2023 Jul;18(7):1463-1471. doi: 10.4103/1673-5374.361544. Neural Regen Res. 2023. PMID: 36571343 Free PMC article. Review.
-
Mono-UFMylation promotes misfolding-associated secretion of α-synuclein.Sci Adv. 2024 Mar 15;10(11):eadk2542. doi: 10.1126/sciadv.adk2542. Epub 2024 Mar 15. Sci Adv. 2024. PMID: 38489364 Free PMC article.
-
Adult-onset neuronal ceroid lipofuscinosis misdiagnosed as autoimmune encephalitis and normal-pressure hydrocephalus: A 10-year case report and case-based review.Medicine (Baltimore). 2024 Oct 25;103(43):e40248. doi: 10.1097/MD.0000000000040248. Medicine (Baltimore). 2024. PMID: 39470529 Free PMC article. Review.
-
Intercellular transmission of alpha-synuclein.Front Mol Neurosci. 2024 Sep 11;17:1470171. doi: 10.3389/fnmol.2024.1470171. eCollection 2024. Front Mol Neurosci. 2024. PMID: 39324117 Free PMC article. Review.
-
Navigating the Gene Co-Expression Network and Drug Repurposing Opportunities for Brain Disorders Associated with Neurocognitive Impairment.Brain Sci. 2023 Nov 7;13(11):1564. doi: 10.3390/brainsci13111564. Brain Sci. 2023. PMID: 38002524 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous