

Inversion of a topological domain leads to restricted changes in its gene expression and affects interdomain communication
- PMID: 35502750
- PMCID: PMC9148567
- DOI: 10.1242/dev.200568
Inversion of a topological domain leads to restricted changes in its gene expression and affects interdomain communication
Abstract
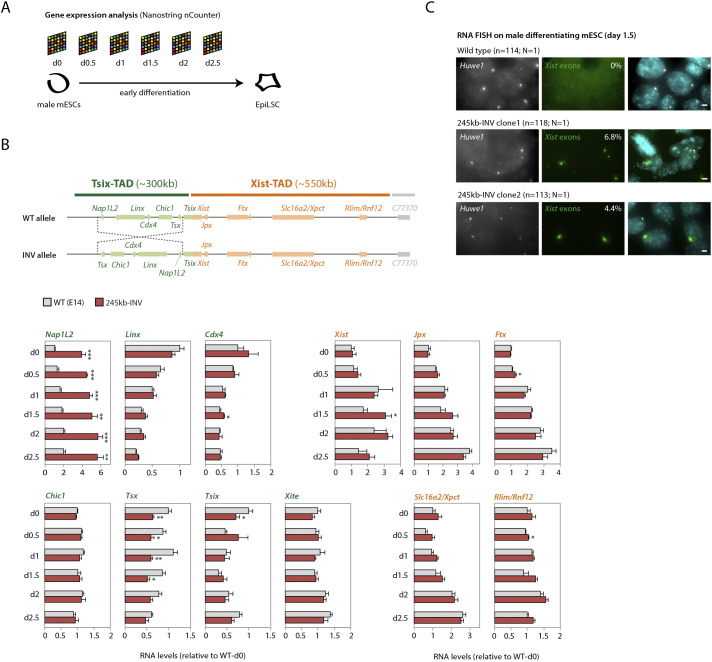
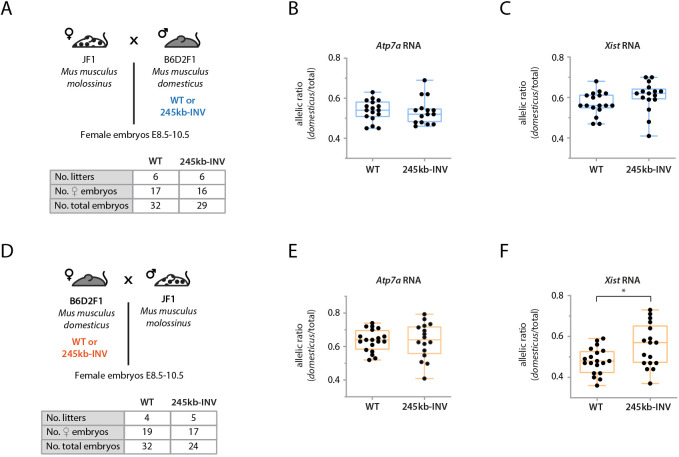
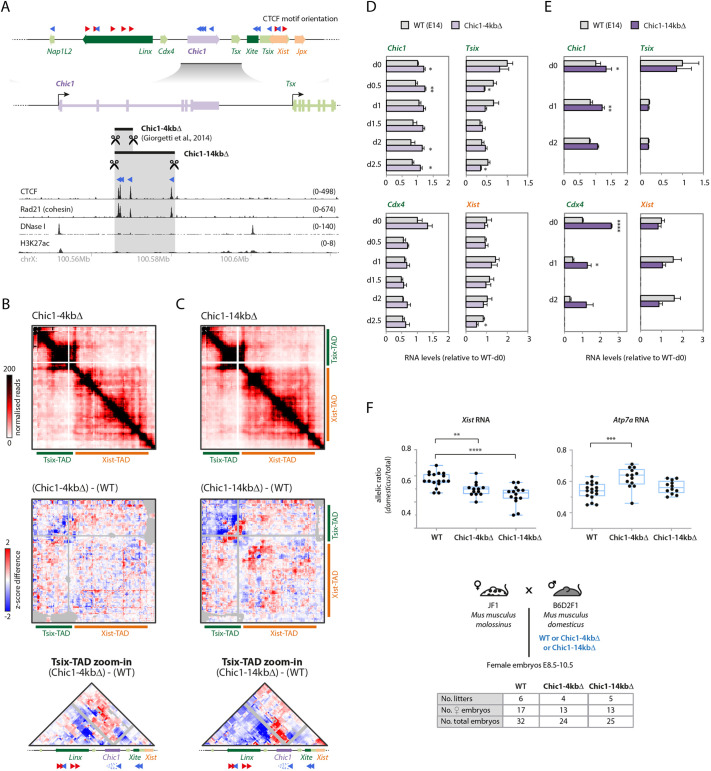
The interplay between the topological organization of the genome and the regulation of gene expression remains unclear. Depletion of molecular factors (e.g. CTCF) underlying topologically associating domains (TADs) leads to modest alterations in gene expression, whereas genomic rearrangements involving TAD boundaries disrupt normal gene expression and can lead to pathological phenotypes. Here, we targeted the TAD neighboring that of the noncoding transcript Xist, which controls X-chromosome inactivation. Inverting 245 kb within the TAD led to expected rearrangement of CTCF-based contacts but revealed heterogeneity in the 'contact' potential of different CTCF sites. Expression of most genes therein remained unaffected in mouse embryonic stem cells and during differentiation. Interestingly, expression of Xist was ectopically upregulated. The same inversion in mouse embryos led to biased Xist expression. Smaller inversions and deletions of CTCF clusters led to similar results: rearrangement of contacts and limited changes in local gene expression, but significant changes in Xist expression in embryos. Our study suggests that the wiring of regulatory interactions within a TAD can influence the expression of genes in neighboring TADs, highlighting the existence of mechanisms of inter-TAD communication.
Keywords: Gene expression; Genomic engineering; TADs; X-inactivation; Xist.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures



Similar articles
-
Structural organization of the inactive X chromosome in the mouse.Nature. 2016 Jul 28;535(7613):575-9. doi: 10.1038/nature18589. Epub 2016 Jul 18. Nature. 2016. PMID: 27437574 Free PMC article.
-
The bipartite TAD organization of the X-inactivation center ensures opposing developmental regulation of Tsix and Xist.Nat Genet. 2019 Jun;51(6):1024-1034. doi: 10.1038/s41588-019-0412-0. Epub 2019 May 27. Nat Genet. 2019. PMID: 31133748 Free PMC article.
-
Clustered CTCF binding is an evolutionary mechanism to maintain topologically associating domains.Genome Biol. 2020 Jan 7;21(1):5. doi: 10.1186/s13059-019-1894-x. Genome Biol. 2020. PMID: 31910870 Free PMC article.
-
Multifaceted role of CTCF in X-chromosome inactivation.Chromosoma. 2024 Oct;133(4):217-231. doi: 10.1007/s00412-024-00826-w. Epub 2024 Oct 21. Chromosoma. 2024. PMID: 39433641 Review.
-
Differential 3D chromatin organization and gene activity in genomic imprinting.Curr Opin Genet Dev. 2020 Apr;61:17-24. doi: 10.1016/j.gde.2020.03.004. Epub 2020 Apr 13. Curr Opin Genet Dev. 2020. PMID: 32299027 Review.
Cited by
-
Orchestrating Asymmetric Expression: Mechanisms behind Xist Regulation.Epigenomes. 2024 Feb 1;8(1):6. doi: 10.3390/epigenomes8010006. Epigenomes. 2024. PMID: 38390897 Free PMC article. Review.
-
invdup(8)(8q24.13q24.3)-A Complex Alteration and Its Clinical Consequences.Genes (Basel). 2024 Jul 12;15(7):910. doi: 10.3390/genes15070910. Genes (Basel). 2024. PMID: 39062689 Free PMC article. Review.
-
Proximity-dependent recruitment of Polycomb repressive complexes by the lncRNA Airn.Cell Rep. 2023 Jul 25;42(7):112803. doi: 10.1016/j.celrep.2023.112803. Epub 2023 Jul 11. Cell Rep. 2023. PMID: 37436897 Free PMC article.
-
Outward-oriented sites within clustered CTCF boundaries are key for intra-TAD chromatin interactions and gene regulation.Nat Commun. 2023 Dec 7;14(1):8101. doi: 10.1038/s41467-023-43849-0. Nat Commun. 2023. PMID: 38062010 Free PMC article.
-
Regulatory principles and mechanisms governing the onset of random X-chromosome inactivation.Curr Opin Genet Dev. 2023 Aug;81:102063. doi: 10.1016/j.gde.2023.102063. Epub 2023 Jun 23. Curr Opin Genet Dev. 2023. PMID: 37356341 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
