

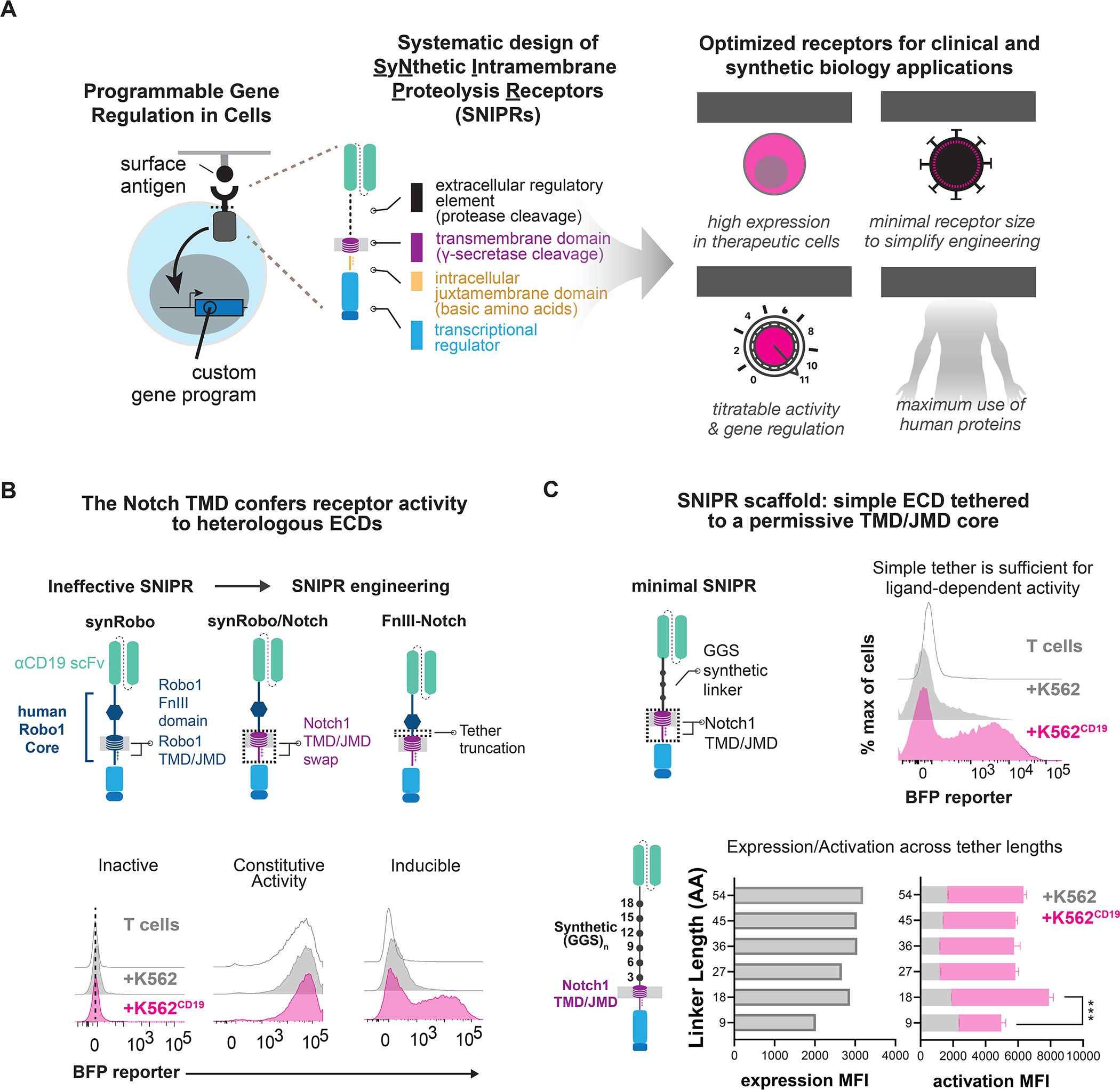
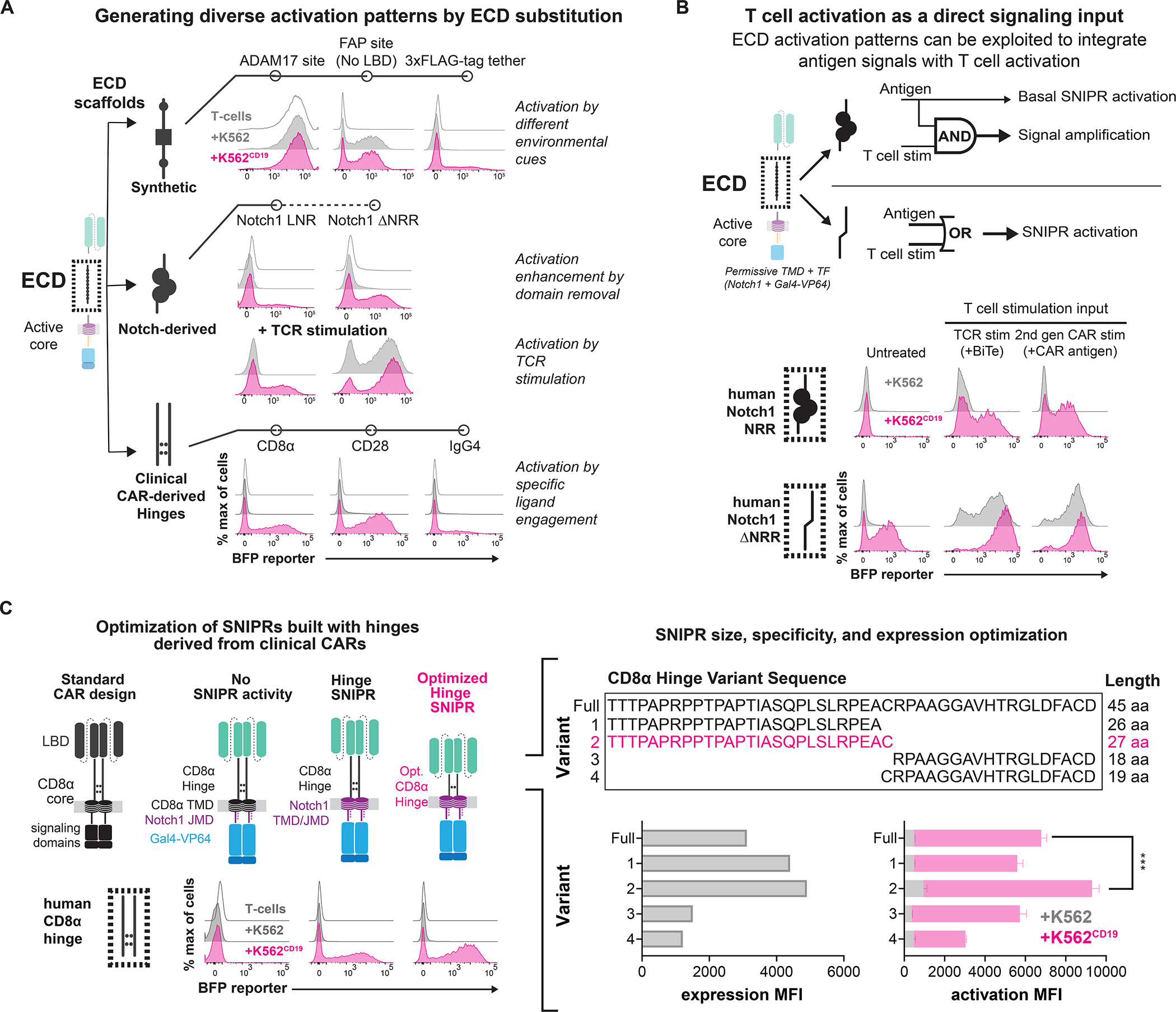
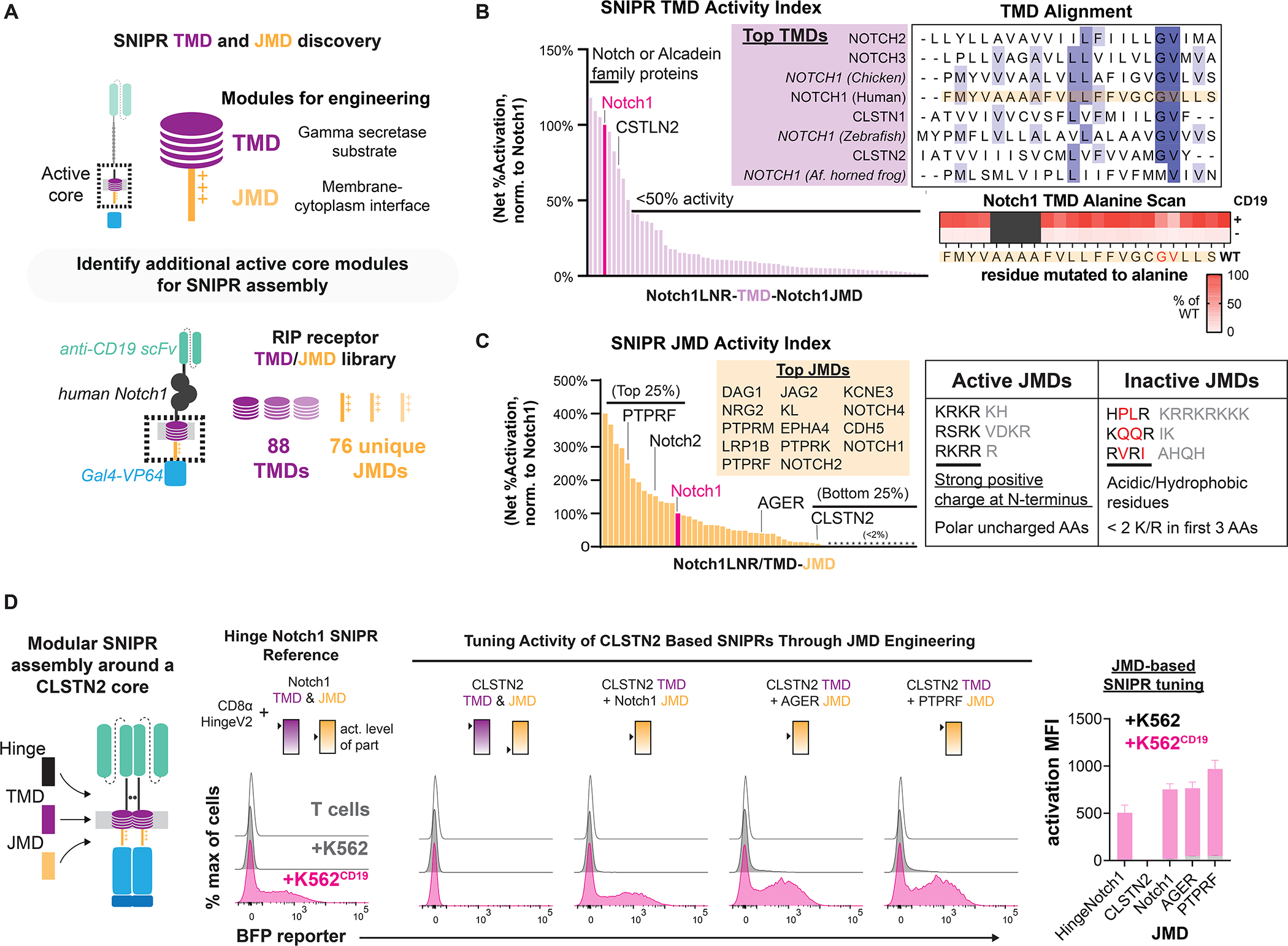
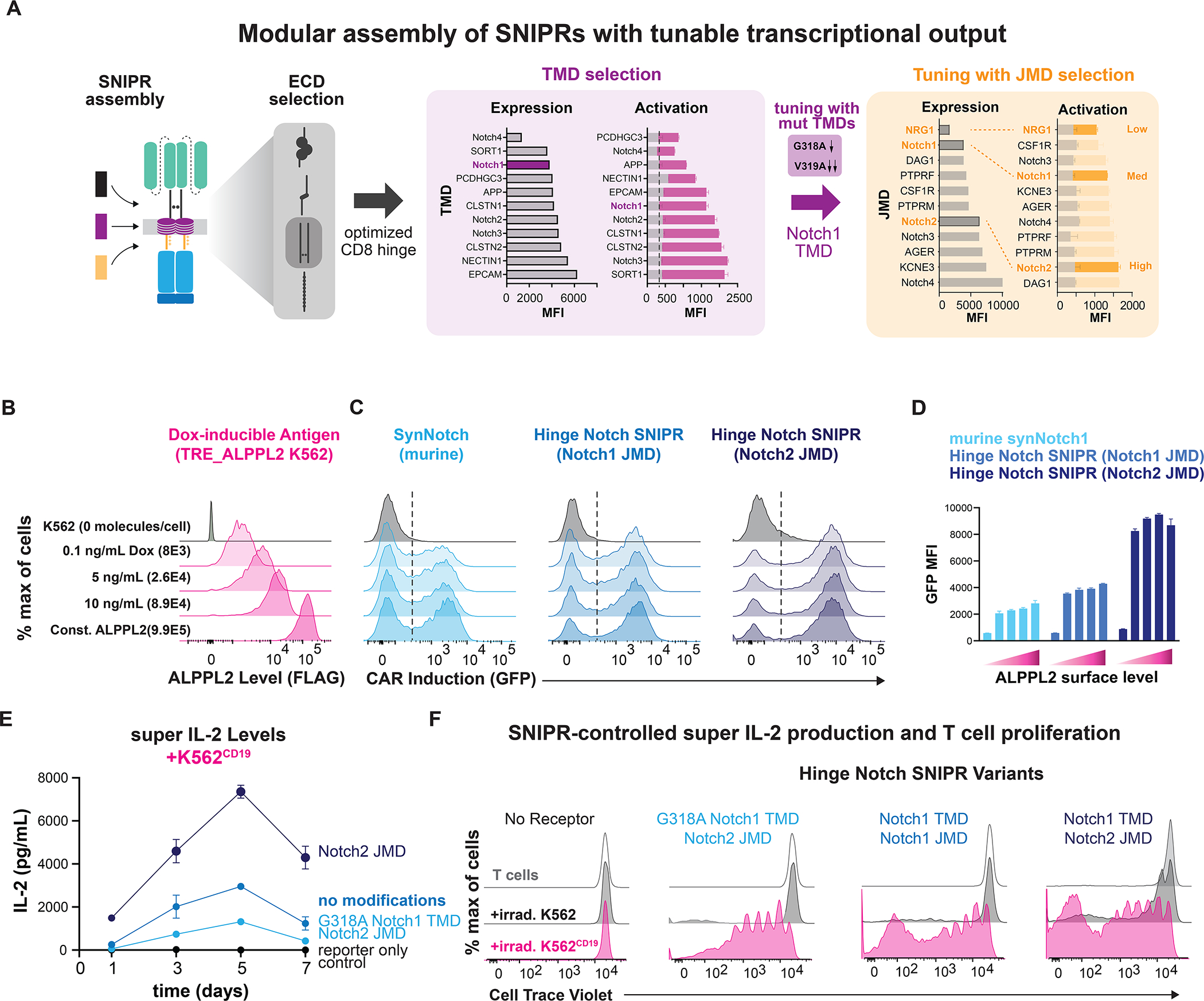
Modular design of synthetic receptors for programmed gene regulation in cell therapies
- PMID: 35427499
- PMCID: PMC9108009
- DOI: 10.1016/j.cell.2022.03.023
Modular design of synthetic receptors for programmed gene regulation in cell therapies
Abstract
Synthetic biology has established powerful tools to precisely control cell function. Engineering these systems to meet clinical requirements has enormous medical implications. Here, we adopted a clinically driven design process to build receptors for the autonomous control of therapeutic cells. We examined the function of key domains involved in regulated intramembrane proteolysis and showed that systematic modular engineering can generate a class of receptors that we call synthetic intramembrane proteolysis receptors (SNIPRs) that have tunable sensing and transcriptional response abilities. We demonstrate the therapeutic potential of the receptor platform by engineering human primary T cells for multi-antigen recognition and production of dosed, bioactive payloads relevant to the treatment of disease. Our design framework enables the development of fully humanized and customizable transcriptional receptors for the programming of therapeutic cells suitable for clinical translation.
Keywords: CAR-T cells; cancer immunotherapy; cell therapy; synNotch; synthetic biology.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests I.Z., R.L., and K.T.R. are co-inventors on patents for synthetic receptors (PRV 62/905,258, 62/905,262, 62/905,266, 62/905,268, 62/905,251, and 62/905,263). R.L. and K.T.R. are co-inventors on patents for synthetic receptors PRV 62/007,807. R.L., I.Z., D.I.P., D.V.I., A.S.K., and K.T.R. are co-inventors for synthetic receptors PRV 63/007,795. K.T.R. is a co-founder of Arsenal Biosciences, consultant, SAB member, and stockholder. K.T.R. is an inventor on patents for synthetic Notch receptors (WO2016138034A1 and PRV/2016/62/333,106) and receives licensing fees and royalties. The patents were licensed by Cell Design Labs and are now part of Gilead. He was a founding scientist/consultant and stockholder in Cell Design Labs, now a Gilead company. K.T.R. holds stock in Gilead. K.T.R. is on the SAB of Ziopharm Oncology and an advisor to Venrock. A.S.K. is a scientific advisor for and holds equity in Senti Biosciences and Chroma Medicine and is a co-founder of Fynch Biosciences and K2 Biotechnologies. B.L. is an inventor on patents (WO2017095823A1 and US20180369409A1) held by University of California that cover ALPPL2-targeted anticancer therapy and ALPPL2-targeting antibodies. Unrelated to this work, B.L. is a founder and stockholder of Fortis Therapeutics and Vivace Therapeutics and a consultant for Merck Sharpe & Dohme.
Figures
Comment in
-
Designer protein circuits enable safe cancer immunotherapy.Nature. 2022 Jun;606(7916):868-869. doi: 10.1038/d41586-022-01528-y. Nature. 2022. PMID: 35654967 No abstract available.
Similar articles
-
Synthetic Immunotherapy: Programming Immune Cells with Novel and Sophisticated Logic Capabilities.Transplant Cell Ther. 2022 Sep;28(9):560-571. doi: 10.1016/j.jtct.2022.06.001. Epub 2022 Jun 10. Transplant Cell Ther. 2022. PMID: 35691572 Review.
-
Engineering T Cells with Customized Therapeutic Response Programs Using Synthetic Notch Receptors.Cell. 2016 Oct 6;167(2):419-432.e16. doi: 10.1016/j.cell.2016.09.011. Epub 2016 Sep 29. Cell. 2016. PMID: 27693353 Free PMC article.
-
Programmable synthetic receptors: the next-generation of cell and gene therapies.Signal Transduct Target Ther. 2024 Jan 3;9(1):7. doi: 10.1038/s41392-023-01680-5. Signal Transduct Target Ther. 2024. PMID: 38167329 Free PMC article. Review.
-
Engineering Axl specific CAR and SynNotch receptor for cancer therapy.Sci Rep. 2018 Mar 1;8(1):3846. doi: 10.1038/s41598-018-22252-6. Sci Rep. 2018. PMID: 29497107 Free PMC article.
-
Molecular Design, Optimization, and Genomic Integration of Chimeric B Cell Receptors in Murine B Cells.Front Immunol. 2019 Nov 14;10:2630. doi: 10.3389/fimmu.2019.02630. eCollection 2019. Front Immunol. 2019. PMID: 31798579 Free PMC article.
Cited by
-
Engineering programmable material-to-cell pathways via synthetic notch receptors to spatially control differentiation in multicellular constructs.Nat Commun. 2024 Jul 13;15(1):5891. doi: 10.1038/s41467-024-50126-1. Nat Commun. 2024. PMID: 39003263 Free PMC article.
-
New tricks for an old pathway: emerging Notch-based biotechnologies and therapeutics.Trends Pharmacol Sci. 2023 Dec;44(12):934-948. doi: 10.1016/j.tips.2023.09.011. Epub 2023 Oct 25. Trends Pharmacol Sci. 2023. PMID: 37891017 Free PMC article. Review.
-
Cooperative assembly confers regulatory specificity and long-term genetic circuit stability.Cell. 2023 Aug 31;186(18):3810-3825.e18. doi: 10.1016/j.cell.2023.07.012. Epub 2023 Aug 7. Cell. 2023. PMID: 37552983 Free PMC article.
-
Adult tissue-specific stem cell interaction: novel technologies and research advances.Front Cell Dev Biol. 2023 Sep 21;11:1220694. doi: 10.3389/fcell.2023.1220694. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37808078 Free PMC article. Review.
-
Mechanisms of Resistance and Treatment of Relapse after CAR T-cell Therapy for Large B-cell Lymphoma and Multiple Myeloma.Transplant Cell Ther. 2023 Jul;29(7):418-428. doi: 10.1016/j.jtct.2023.04.007. Epub 2023 Apr 17. Transplant Cell Ther. 2023. PMID: 37076102 Free PMC article. Review.
References
-
- Brou C, Logeat F, Gupta N, Bessia C, LeBail O, Doedens JR, Cumano A, Roux P, Black RA, and Israël A (2000). A novel proteolytic cleavage involved in Notch signaling: the role of the disintegrin-metalloprotease TACE. Mol Cell 5, 207–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
