

Review
doi: 10.3390/jcdd9030073.
In Vivo Methods to Monitor Cardiomyocyte Proliferation
Affiliations
- PMID: 35323621
- PMCID: PMC8950582
- DOI: 10.3390/jcdd9030073
Item in Clipboard
Review
In Vivo Methods to Monitor Cardiomyocyte Proliferation
J Cardiovasc Dev Dis.
.
Abstract
Adult mammalian cardiomyocytes demonstrate scarce cycling and even lower proliferation rates in response to injury. Signals that enhance cardiomyocyte proliferation after injury will be groundbreaking, address unmet clinical needs, and represent new strategies to treat cardiovascular diseases. In vivo methods to monitor cardiomyocyte proliferation are critical to addressing this challenge. Fortunately, advances in transgenic approaches provide sophisticated techniques to quantify cardiomyocyte cycling and proliferation.
Keywords: cardiomyocyte cycling; heart; mouse; regeneration; transgenics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
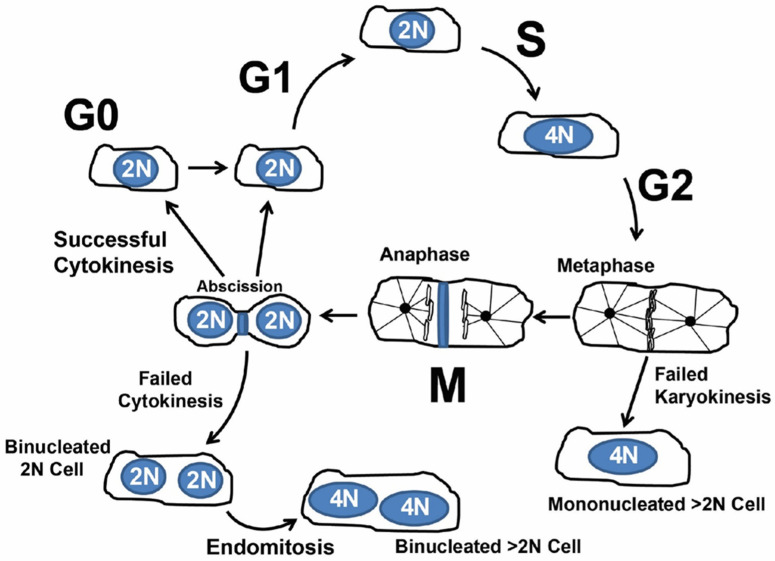
Simplified overview of cardiomyocyte cycling. After G2, failed karyokinesis (nuclear division) produces a single nucleus with >2N genomic DNA content. After mitosis and karyokinesis failed cytokinesis produces a binucleated cell. Further endomitosis of binucleated (on multinucleated) cells produces nuclei with >2N genomic content.
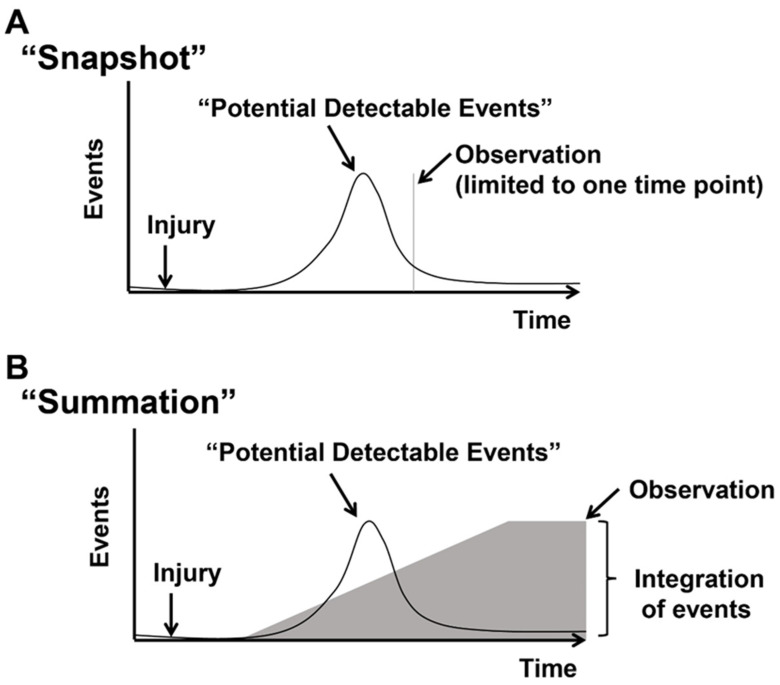
Detecting “Actively cycling” and “Previously cycled” cells. Events represent cycling cells. After an injury, there is an increase and decrease in cycling cells. (A) The “snapshot” approach measures cycling events in blue at a distinct time point after the injury. Typically, cycling cells are detected by the co-localization of markers of cycling (Ki67, PH3, Aurora B, and Anillin) with cardiomyocytes or transgenic mice that report phases of the cell cycle (FUCCI, Ki67-RFP, eGFP-Anillin). (B) The “summation” approach relies on indelibly marking cycling events and measuring total events after an injury. Cycled cells are labeled by incorporating nucleotide analogs into synthesizing genomic DNA (3H-Thymidine, Bromodeoxyuridine, or 5-Ethynyl-2-deoxyuridine) or transgenic mice that mark cells that cycled or underwent mitosis (MADM, Confetti/Brainbow, and αDKRC::RLTG). Figure adapted from Bradley et al., Circ Res 2021, 128, 155–168.
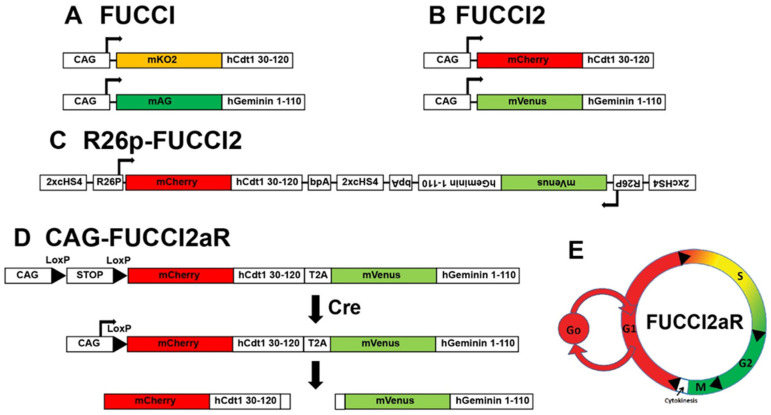
Fluorescent Ubiquitination-based Cell Cycle Indicator (FUCCI) reporters. First-generation FUCCI (A) and FUCCI2 (B). Second-generation Bidirectional R26p-FUCCI2 (C) and polycistronic FUCCI2aR (D). (E) Schematic of cell cycle phase-dependent differential expression of FUCCIs.
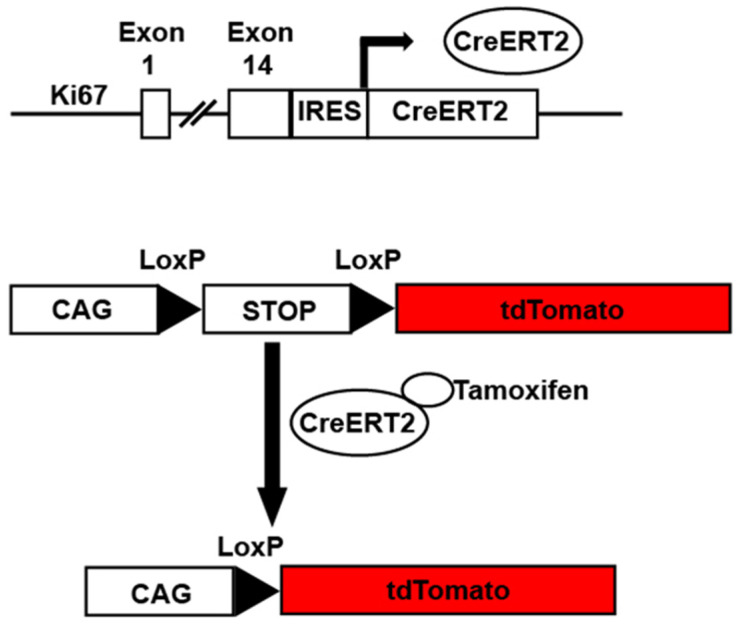
Ki67iresCreER mice. Example of Ki67iresCreER::CAG-Lox-STOP-Lox-tdTomato mice. In the presence of tamoxifen, cells undergoing cycling as defined by Ki67 expression have activated Cre recombinase. The activated Cre excises a STOP cassette allowing tdTomato expression under the control of the constitutively active CAG promoter.
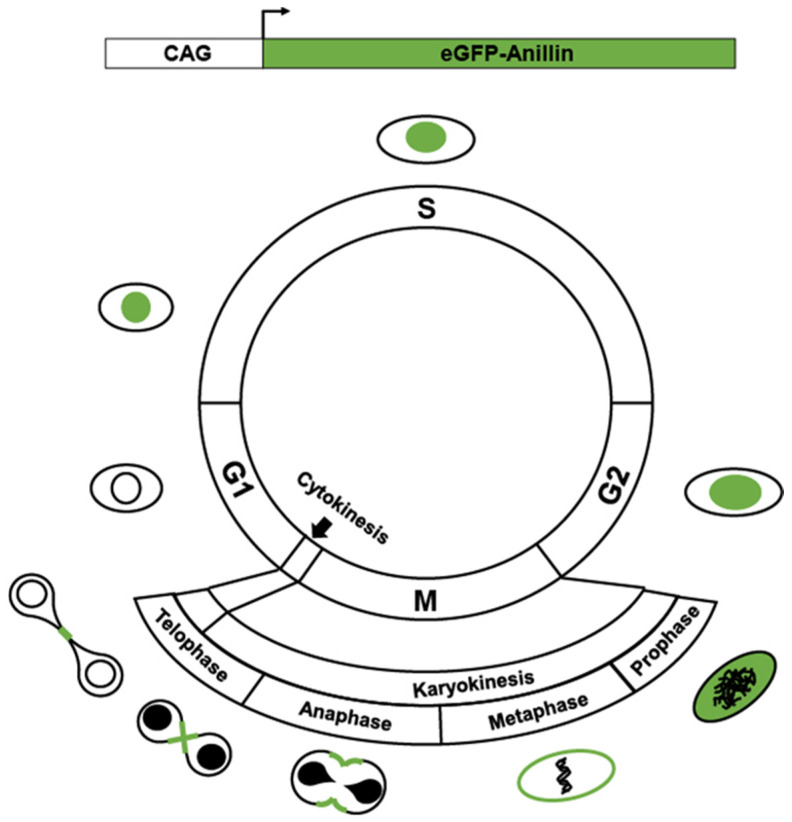
CAG-eGFP-Anillin. The subcellular localization of constitutive expression of eGFP-Anillin marks cells that undergo cytokinesis by labeling the mid-body. Figure adapted from Hesse, M. et al., Nat Commun 2012, 3, 1076.
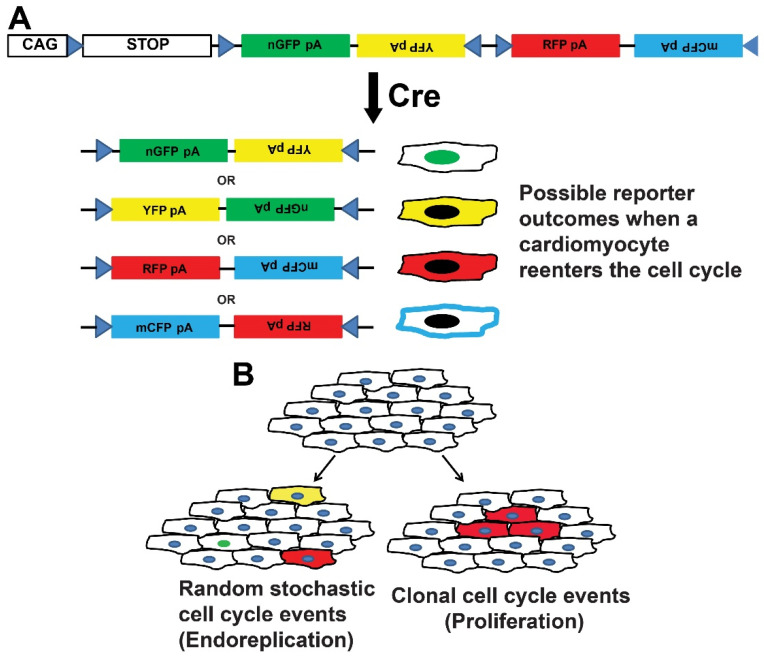
CAG-loxP-STOP-loxP-Confetti (Brainbow). (A) The tamoxifen-induced expression of Cre-dependent causes stochastic recombination of a multi-fluorescent reporter. Identifying of clusters of cells expressing the same fluorescent reporters are used to infer clonal expansion. (B) Schematic of the interpretation of Confetti expression used to identify random stochastic events (left) and clonal expansion consistent with proliferation (right).
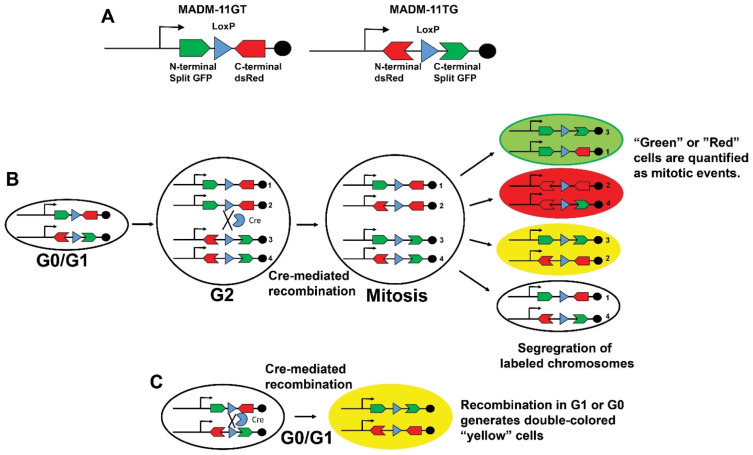
Mosaic Analysis with Double Markers (MADM). (A) Schematic of MADM-11GT and MADM-11TG transgenes that harbor spilt GFP and dsRed constructs with intronic LoxP sites. (B) Tamoxifen exposure induces the Cre recombinase expression that catalyzes an inter-chromosomal rearrangement of split fluorescent reporters and labels mitotic events. (C) Recombination during G1 or G1 produces double colored “Yellow” cells. The detection of “Green” or “Red” cells in panel (B) allows the quantification of mitotic events. Figure adapted from Zong, H. et al., Cell 2005, 121, 479–492.
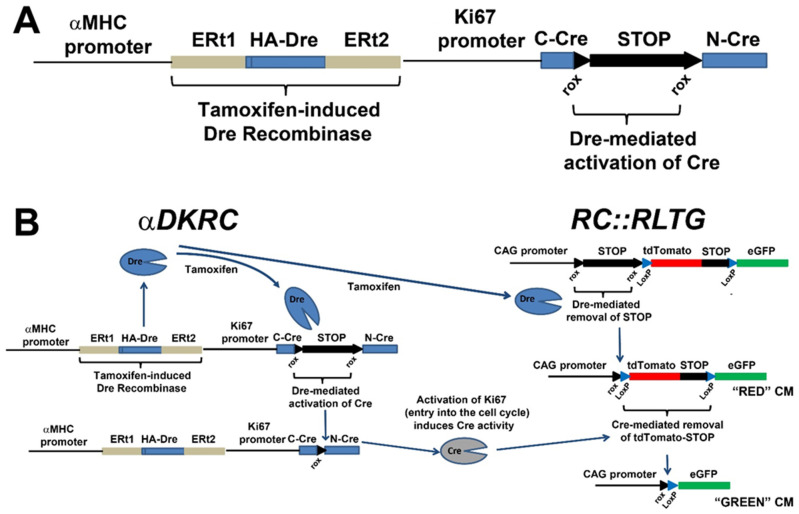
αDKRC::RLTG. (A) The αDKRC transgene. (B) Schematic outlining the expression of GFP in adult cycling cardiomyocytes. Tamoxifen exposure induces the cardiomyocyte-specific Dre-recombinase and subsequent excision of derived poly(A) signal repeats and flanked by two loxP sites from the RLTG reporter, resulting in the expression of tdTomato and RoxedCre to generate catalytically active Cre recombinase under control to the Ki67 cell cycle promoter. Cardiomyocytes reentering the cell cycle, as defined by activation of the Ki67 promoter, express Cre and excise the tdTomato-STOP cassette resulting in the expression of eGFP (enhanced green fluorescent proteins) in cardiomyocytes. A short duration of Tamoxifen exposure activates that reporter system. Schematic of expected changes in tdTomato and eGFP-labeled cardiomyocytes. Figure adapted from Bradley et al., Circ Res 2021, 128, 155–168.
Similar articles
-
Long Noncoding RNA CPR (Cardiomyocyte Proliferation Regulator) Regulates Cardiomyocyte Proliferation and Cardiac Repair.Circulation. 2019 Jun 4;139(23):2668-2684. doi: 10.1161/CIRCULATIONAHA.118.035832. Epub 2019 Mar 5. Circulation. 2019. PMID: 30832495
-
Stereological estimation of cardiomyocyte number and proliferation.Methods. 2021 Jun;190:55-62. doi: 10.1016/j.ymeth.2020.06.002. Epub 2020 Jun 27. Methods. 2021. PMID: 32603825
-
Cardiomyocyte cell cycle dynamics and proliferation revealed through cardiac-specific transgenesis of fluorescent ubiquitinated cell cycle indicator (FUCCI).J Mol Cell Cardiol. 2019 Feb;127:154-164. doi: 10.1016/j.yjmcc.2018.12.007. Epub 2018 Dec 18. J Mol Cell Cardiol. 2019. PMID: 30571978 Free PMC article.
-
Towards regenerating the mammalian heart: challenges in evaluating experimentally induced adult mammalian cardiomyocyte proliferation.Am J Physiol Heart Circ Physiol. 2016 May 1;310(9):H1045-54. doi: 10.1152/ajpheart.00697.2015. Epub 2016 Feb 26. Am J Physiol Heart Circ Physiol. 2016. PMID: 26921436 Review.
-
Cardiomyocyte proliferation in cardiac development and regeneration: a guide to methodologies and interpretations.Am J Physiol Heart Circ Physiol. 2015 Oct;309(8):H1237-50. doi: 10.1152/ajpheart.00559.2015. Epub 2015 Sep 4. Am J Physiol Heart Circ Physiol. 2015. PMID: 26342071 Review.
References
-
- Benjamin E.J., Muntner P., Alonso A., Bittencourt M.S., Callaway C.W., Carson A.P., Chamberlain A.M., Chang A.R., Cheng S., Das S.R., et al. Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association. Circulation. 2019;139:e56–e528. doi: 10.1161/CIR.0000000000000659. - DOI - PubMed
-
- Yancy C.W., Jessup M., Bozkurt B., Butler J., Casey D.E., Jr., Drazner M.H., Fonarow G.C., Geraci S.A., Horwich T., Januzzi J.L., et al. 2013 ACCF/AHA guideline for the management of heart failure: Executive summary: A report of the American College of Cardiology Foundation/American Heart Association Task Force on practice guidelines. Circulation. 2013;128:1810–1852. doi: 10.1161/CIR.0b013e31829e8807. - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
