

Subcellular Remodeling in Filamin C Deficient Mouse Hearts Impairs Myocyte Tension Development during Progression of Dilated Cardiomyopathy
- PMID: 35055055
- PMCID: PMC8779483
- DOI: 10.3390/ijms23020871
Subcellular Remodeling in Filamin C Deficient Mouse Hearts Impairs Myocyte Tension Development during Progression of Dilated Cardiomyopathy
Abstract
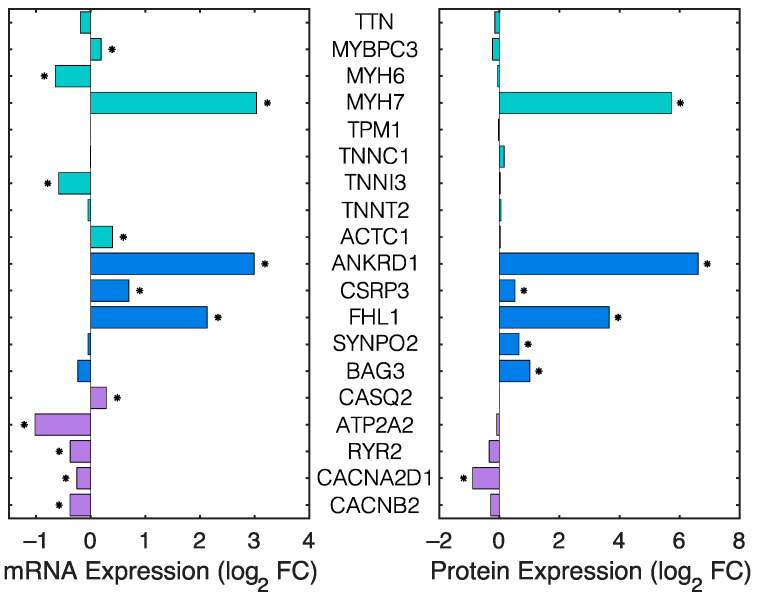
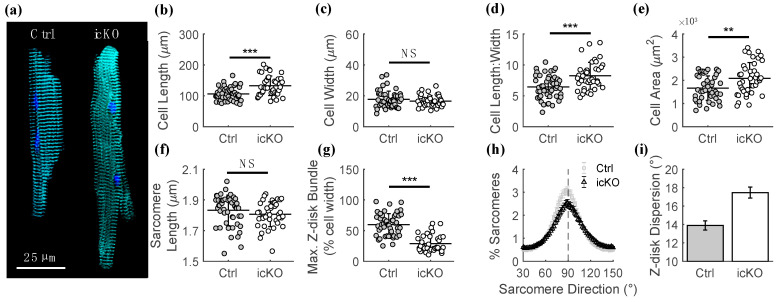
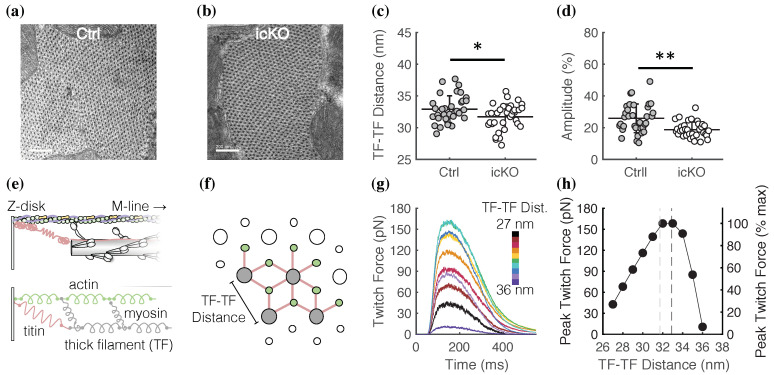
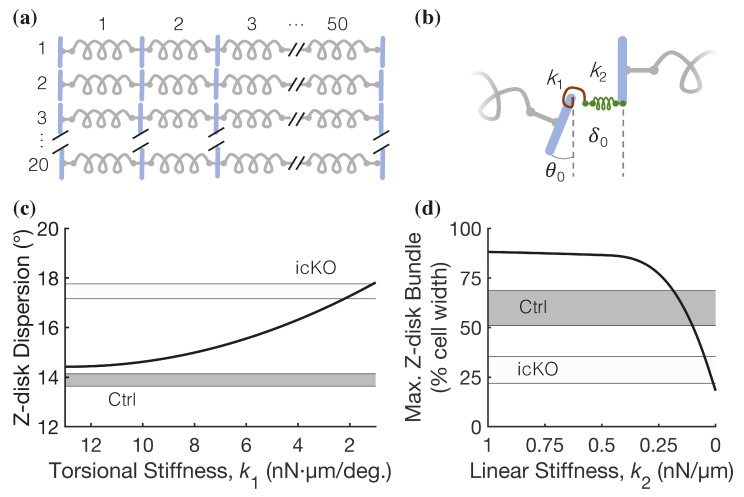
Dilated cardiomyopathy (DCM) is a life-threatening form of heart disease that is typically characterized by progressive thinning of the ventricular walls, chamber dilation, and systolic dysfunction. Multiple mutations in the gene encoding filamin C (FLNC), an actin-binding cytoskeletal protein in cardiomyocytes, have been found in patients with DCM. However, the mechanisms that lead to contractile impairment and DCM in patients with FLNC variants are poorly understood. To determine how FLNC regulates systolic force transmission and DCM remodeling, we used an inducible, cardiac-specific FLNC-knockout (icKO) model to produce a rapid onset of DCM in adult mice. Loss of FLNC reduced systolic force development in single cardiomyocytes and isolated papillary muscles but did not affect twitch kinetics or calcium transients. Electron and immunofluorescence microscopy showed significant defects in Z-disk alignment in icKO mice and altered myofilament lattice geometry. Moreover, a loss of FLNC induces a softening myocyte cortex and structural adaptations at the subcellular level that contribute to disrupted longitudinal force production during contraction. Spatially explicit computational models showed that these structural defects could be explained by a loss of inter-myofibril elastic coupling at the Z-disk. Our work identifies FLNC as a key regulator of the multiscale ultrastructure of cardiomyocytes and therefore plays an important role in maintaining systolic mechanotransmission pathways, the dysfunction of which may be key in driving progressive DCM.
Keywords: Z-disk; cardiac muscle; cellular remodeling; computational modeling; mechanotransmission; sarcomere.
Conflict of interest statement
A.D.M. is a co-founder of and has an equity interest in Insilicomed Inc. and an equity interest in Vektor Medical, Inc. He serves on the scientific advisory board of Insilicomed, and as a scientific advisor to both companies. Some of his research grants have been identified for conflict-of-interest management based on the overall scope of the project and its potential benefit to these companies. The author is required to disclose this relationship in publications acknowledging the grant support; however, the research subject and findings reported in this study did not involve the companies in any way and have no relationship with the business activities or scientific interests of either company. The terms of this arrangement have been reviewed and approved by the University of California San Diego in accordance with its conflict-of-interest policies.
Figures


Similar articles
-
Interaction of Filamin C With Actin Is Essential for Cardiac Development and Function.Circ Res. 2023 Aug 18;133(5):400-411. doi: 10.1161/CIRCRESAHA.123.322750. Epub 2023 Jul 26. Circ Res. 2023. PMID: 37492967 Free PMC article.
-
Filamin C Truncation Mutations Are Associated With Arrhythmogenic Dilated Cardiomyopathy and Changes in the Cell-Cell Adhesion Structures.JACC Clin Electrophysiol. 2018 Apr;4(4):504-514. doi: 10.1016/j.jacep.2017.12.003. Epub 2018 Feb 2. JACC Clin Electrophysiol. 2018. PMID: 30067491 Free PMC article.
-
Modulating the tension-time integral of the cardiac twitch prevents dilated cardiomyopathy in murine hearts.JCI Insight. 2020 Oct 15;5(20):e142446. doi: 10.1172/jci.insight.142446. JCI Insight. 2020. PMID: 32931484 Free PMC article.
-
Filamin C in cardiomyopathy: from physiological roles to DNA variants.Heart Fail Rev. 2022 Jul;27(4):1373-1385. doi: 10.1007/s10741-021-10172-z. Epub 2021 Sep 17. Heart Fail Rev. 2022. PMID: 34535832 Review.
-
A mutation update for the FLNC gene in myopathies and cardiomyopathies.Hum Mutat. 2020 Jun;41(6):1091-1111. doi: 10.1002/humu.24004. Epub 2020 Mar 20. Hum Mutat. 2020. PMID: 32112656 Free PMC article. Review.
Cited by
-
Nuclear mechanosignaling in striated muscle diseases.Front Physiol. 2023 Mar 7;14:1126111. doi: 10.3389/fphys.2023.1126111. eCollection 2023. Front Physiol. 2023. PMID: 36960155 Free PMC article. Review.
-
Comprehensive review on gene mutations contributing to dilated cardiomyopathy.Front Cardiovasc Med. 2023 Dec 1;10:1296389. doi: 10.3389/fcvm.2023.1296389. eCollection 2023. Front Cardiovasc Med. 2023. PMID: 38107262 Free PMC article. Review.
-
Force-Dependent Structural Changes of Filamin C Rod Domains Regulated by Filamin C Dimer.J Am Chem Soc. 2023 Jul 12;145(27):14670-14678. doi: 10.1021/jacs.3c02303. Epub 2023 Jun 27. J Am Chem Soc. 2023. PMID: 37369984 Free PMC article.
-
Generation of two induced pluripotent stem cell lines from dilated cardiomyopathy patients carrying heterozygous FLNC mutations.Stem Cell Res. 2022 Oct;64:102928. doi: 10.1016/j.scr.2022.102928. Epub 2022 Sep 26. Stem Cell Res. 2022. PMID: 36194907 Free PMC article.
-
Structural and signaling proteins in the Z-disk and their role in cardiomyopathies.Front Physiol. 2023 Mar 2;14:1143858. doi: 10.3389/fphys.2023.1143858. eCollection 2023. Front Physiol. 2023. PMID: 36935760 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
- R01 HL144872/HL/NHLBI NIH HHS/United States
- P30 AR074990/AR/NIAMS NIH HHS/United States
- R01 HL157115/HL/NHLBI NIH HHS/United States
- R01 HL153032/HL/NHLBI NIH HHS/United States
- K99 HL159224/HL/NHLBI NIH HHS/United States
- R01 HL158981/HL/NHLBI NIH HHS/United States
- T32 HL007444/HL/NHLBI NIH HHS/United States
- R01 HL155826/HL/NHLBI NIH HHS/United States
- R01 HL121754/HL/NHLBI NIH HHS/United States
- R01 HL146759/HL/NHLBI NIH HHS/United States
- R00 HL143210/HL/NHLBI NIH HHS/United States
- F32 HL152573/HL/NHLBI NIH HHS/United States
- R01 HL158761/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
