

Celebrating 20 years of live single-actin-filament studies with five golden rules
- PMID: 35042781
- PMCID: PMC8784122
- DOI: 10.1073/pnas.2109506119
Celebrating 20 years of live single-actin-filament studies with five golden rules
Abstract
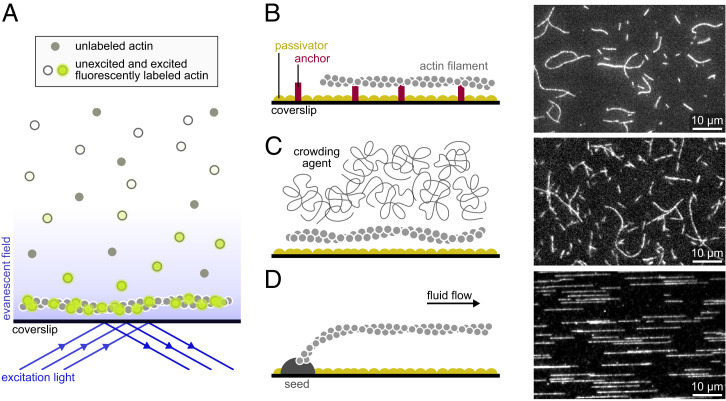
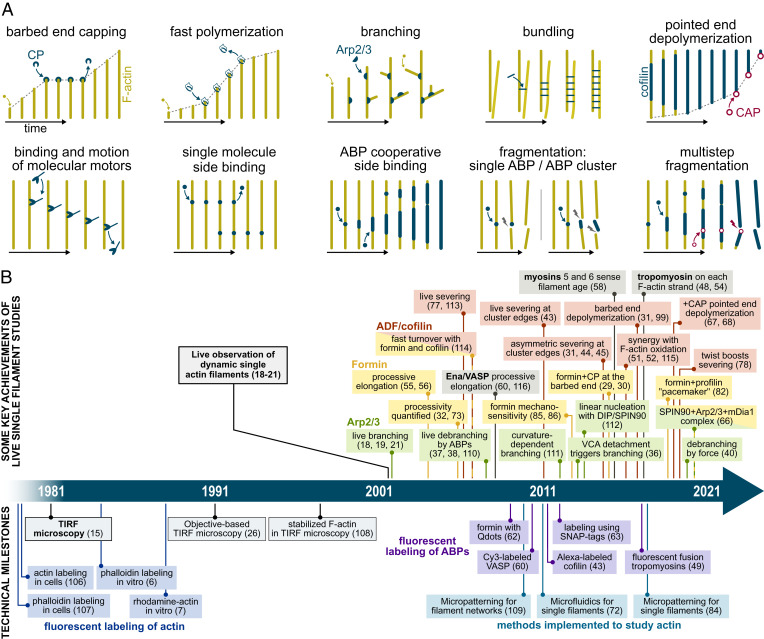
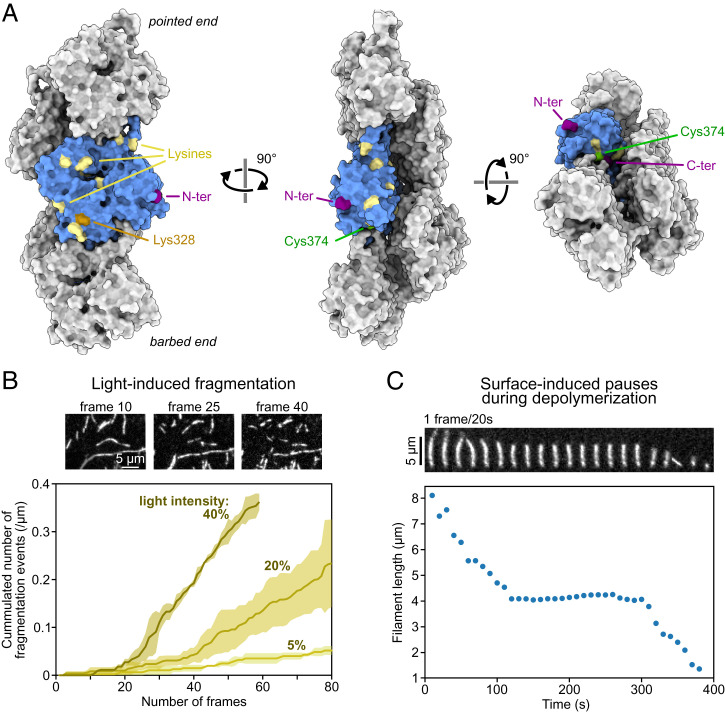
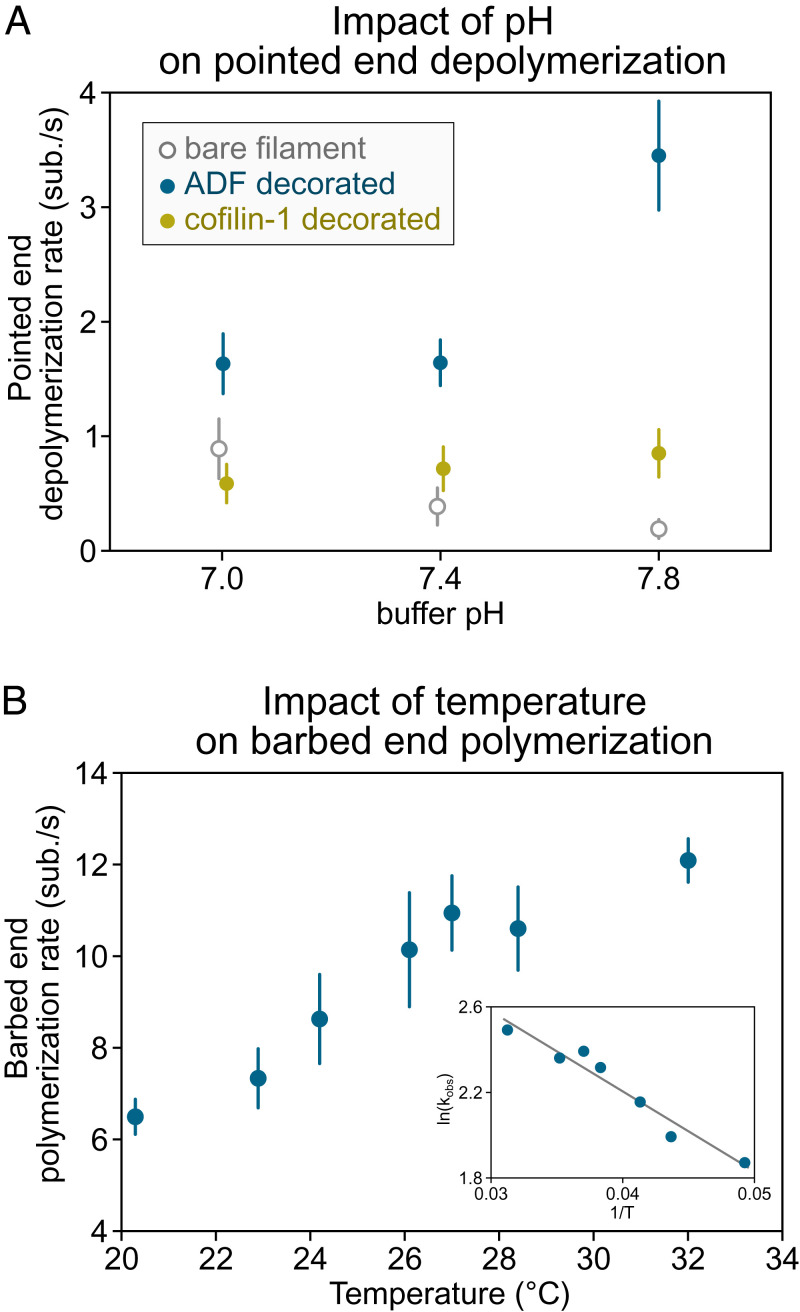
The precise assembly and disassembly of actin filaments is required for several cellular processes, and their regulation has been scrutinized for decades. Twenty years ago, a handful of studies marked the advent of a new type of experiment to study actin dynamics: using optical microscopy to look at individual events, taking place on individual filaments in real time. Here, we summarize the main characteristics of this approach and how it has changed our ability to understand actin assembly dynamics. We also highlight some of its caveats and reflect on what we have learned over the past 20 years, leading us to propose a set of guidelines, which we hope will contribute to a better exploitation of this powerful tool.
Keywords: TIRF; biochemistry; biophysics; cytoskeleton; microscopy.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Biophysics of actin filament severing by cofilin.FEBS Lett. 2013 Apr 17;587(8):1215-9. doi: 10.1016/j.febslet.2013.01.062. Epub 2013 Feb 5. FEBS Lett. 2013. PMID: 23395798 Free PMC article. Review.
-
Microfluidics-Assisted TIRF Imaging to Study Single Actin Filament Dynamics.Curr Protoc Cell Biol. 2017 Dec 11;77:12.13.1-12.13.24. doi: 10.1002/cpcb.31. Curr Protoc Cell Biol. 2017. PMID: 29227552 Free PMC article.
-
Single-molecule studies of actin assembly and disassembly factors.Methods Enzymol. 2014;540:95-117. doi: 10.1016/B978-0-12-397924-7.00006-6. Methods Enzymol. 2014. PMID: 24630103 Free PMC article.
-
Mechanisms of actin disassembly.Mol Biol Cell. 2013 Aug;24(15):2299-302. doi: 10.1091/mbc.E12-09-0694. Mol Biol Cell. 2013. PMID: 23900650 Free PMC article.
-
Cytoskeletal tropomyosins: choreographers of actin filament functional diversity.J Muscle Res Cell Motil. 2013 Aug;34(3-4):261-74. doi: 10.1007/s10974-013-9355-8. Epub 2013 Aug 1. J Muscle Res Cell Motil. 2013. PMID: 23904035 Free PMC article. Review.
Cited by
-
Visualizing Actin and Microtubule Coupling Dynamics In Vitro by Total Internal Reflection Fluorescence (TIRF) Microscopy.J Vis Exp. 2022 Jul 20;(185):10.3791/64074. doi: 10.3791/64074. J Vis Exp. 2022. PMID: 35938818 Free PMC article.
-
Computational tools for quantifying actin filament numbers, lengths, and bundling.Biol Open. 2024 Mar 1;13(3):bio060267. doi: 10.1242/bio.060267. Epub 2024 Mar 1. Biol Open. 2024. PMID: 38372564 Free PMC article.
-
Superresolution microscopy reveals actomyosin dynamics in medioapical arrays.Mol Biol Cell. 2022 Sep 15;33(11):ar94. doi: 10.1091/mbc.E21-11-0537. Epub 2022 May 11. Mol Biol Cell. 2022. PMID: 35544300 Free PMC article.
References
-
- Depue R. H. Jr., Rice R. V., F-actin is a right-handed helix. J. Mol. Biol. 12, 302–303 (1965). - PubMed
-
- Kouyama T., Mihashi K., Fluorimetry study of N-(1-pyrenyl)iodoacetamide-labelled F-actin. Local structural change of actin protomer both on polymerization and on binding of heavy meromyosin. Eur. J. Biochem. 114, 33–38 (1981). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
