

VPS32, a member of the ESCRT complex, modulates adherence to host cells in the parasite Trichomonas vaginalis by affecting biogenesis and cargo sorting of released extracellular vesicles
- PMID: 34951683
- PMCID: PMC11073171
- DOI: 10.1007/s00018-021-04083-3
VPS32, a member of the ESCRT complex, modulates adherence to host cells in the parasite Trichomonas vaginalis by affecting biogenesis and cargo sorting of released extracellular vesicles
Abstract
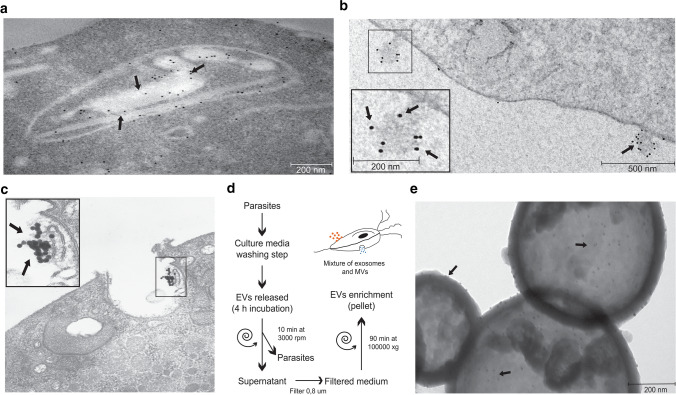
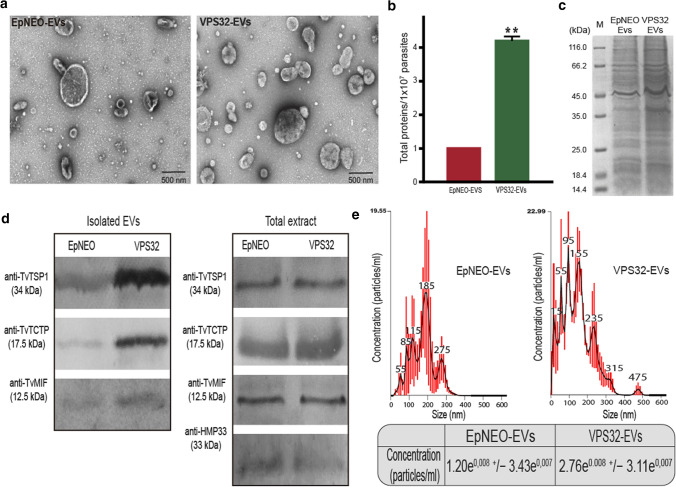
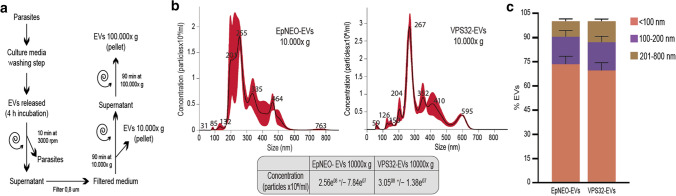
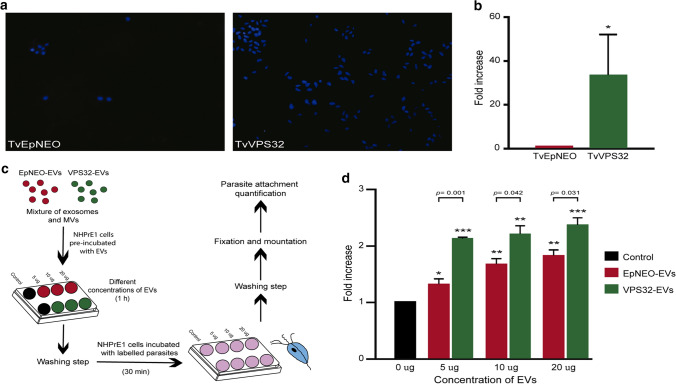
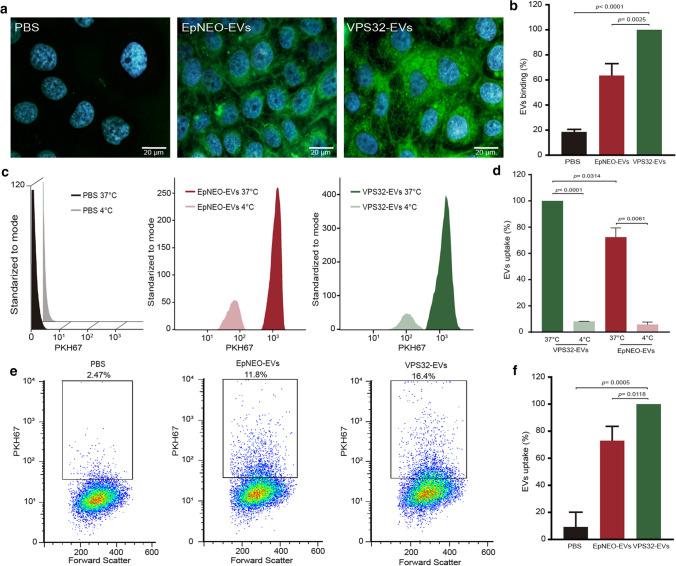
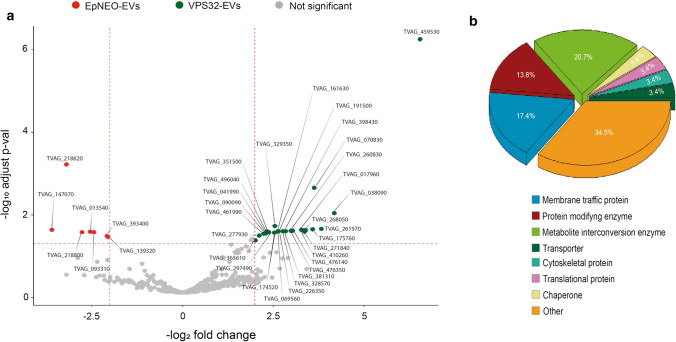
Trichomonas vaginalis is a common sexually transmitted extracellular parasite that adheres to epithelial cells in the human urogenital tract. Extracellular vesicles (EVs) have been described as important players in the pathogenesis of this parasite as they deliver proteins and RNA into host cells and modulate parasite adherence. EVs are heterogeneous membrane vesicles released from virtually all cell types that collectively represent a new dimension of intercellular communication. The Endosomal Sorting Complex Required for Transport (ESCRT) machinery contributes to several key mechanisms in which it reshapes membranes. Based on this, some components of the ESCRT have been implicated in EVs biogenesis in other cells. Here, we demonstrated that VPS32, a member of ESCRTIII complex, contribute to the biogenesis and cargo sorting of extracellular vesicles in the parasite T. vaginalis. Moreover, we observe that parasites overexpressing VPS32 have a striking increase in adherence to host cells compared to control parasites; demonstrating a key role for this protein in mediating host: parasite interactions. These results provide valuable information on the molecular mechanisms involved in extracellular vesicles biogenesis, cargo-sorting, and parasite pathogenesis.
Keywords: ESCRTIII complex; Extracellular vesicles; Trichomonas vaginalis; VPS32.
© 2021. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Similar articles
-
Biogenesis of extracellular vesicles in protozoan parasites: The ESCRT complex in the trafficking fast lane?PLoS Pathog. 2023 Feb 23;19(2):e1011140. doi: 10.1371/journal.ppat.1011140. eCollection 2023 Feb. PLoS Pathog. 2023. PMID: 36821560 Free PMC article. Review.
-
Membrane-shed vesicles from the parasite Trichomonas vaginalis: characterization and their association with cell interaction.Cell Mol Life Sci. 2018 Jun;75(12):2211-2226. doi: 10.1007/s00018-017-2726-3. Epub 2017 Dec 8. Cell Mol Life Sci. 2018. PMID: 29222644 Free PMC article.
-
Trichomonas vaginalis extracellular vesicles up-regulate and directly transfer adherence factors promoting host cell colonization.Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2401159121. doi: 10.1073/pnas.2401159121. Epub 2024 Jun 12. Proc Natl Acad Sci U S A. 2024. PMID: 38865261 Free PMC article.
-
Trichomonas vaginalis exosomes deliver cargo to host cells and mediate host∶parasite interactions.PLoS Pathog. 2013;9(7):e1003482. doi: 10.1371/journal.ppat.1003482. Epub 2013 Jul 11. PLoS Pathog. 2013. PMID: 23853596 Free PMC article.
-
Extracellular vesicles released by anaerobic protozoan parasites: Current situation.Cell Microbiol. 2020 Nov;22(11):e13257. doi: 10.1111/cmi.13257. Epub 2020 Sep 10. Cell Microbiol. 2020. PMID: 32858768 Review.
Cited by
-
An in-depth exploration of the multifaceted roles of EVs in the context of pathogenic single-cell microorganisms.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0003724. doi: 10.1128/mmbr.00037-24. Epub 2024 Jun 13. Microbiol Mol Biol Rev. 2024. PMID: 38869292 Review.
-
Guidelines for the purification and characterization of extracellular vesicles of parasites.J Extracell Biol. 2023 Oct 19;2(10):e117. doi: 10.1002/jex2.117. eCollection 2023 Oct. J Extracell Biol. 2023. PMID: 38939734 Free PMC article.
-
Membranes on the move: The functional role of the extracellular vesicle membrane for contact-dependent cellular signalling.J Extracell Vesicles. 2024 Apr;13(4):e12436. doi: 10.1002/jev2.12436. J Extracell Vesicles. 2024. PMID: 38649339 Free PMC article. Review.
-
Protozoa-Derived Extracellular Vesicles on Intercellular Communication with Special Emphasis on Giardia lamblia.Microorganisms. 2022 Dec 7;10(12):2422. doi: 10.3390/microorganisms10122422. Microorganisms. 2022. PMID: 36557675 Free PMC article. Review.
-
Biogenesis of extracellular vesicles in protozoan parasites: The ESCRT complex in the trafficking fast lane?PLoS Pathog. 2023 Feb 23;19(2):e1011140. doi: 10.1371/journal.ppat.1011140. eCollection 2023 Feb. PLoS Pathog. 2023. PMID: 36821560 Free PMC article. Review.
References
-
- WHO (2018) Report on global sexually transmitted infection surveillance. pp 63
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
