

Crosstalk between Neuron and Glial Cells in Oxidative Injury and Neuroprotection
- PMID: 34948108
- PMCID: PMC8709409
- DOI: 10.3390/ijms222413315
Crosstalk between Neuron and Glial Cells in Oxidative Injury and Neuroprotection
Abstract
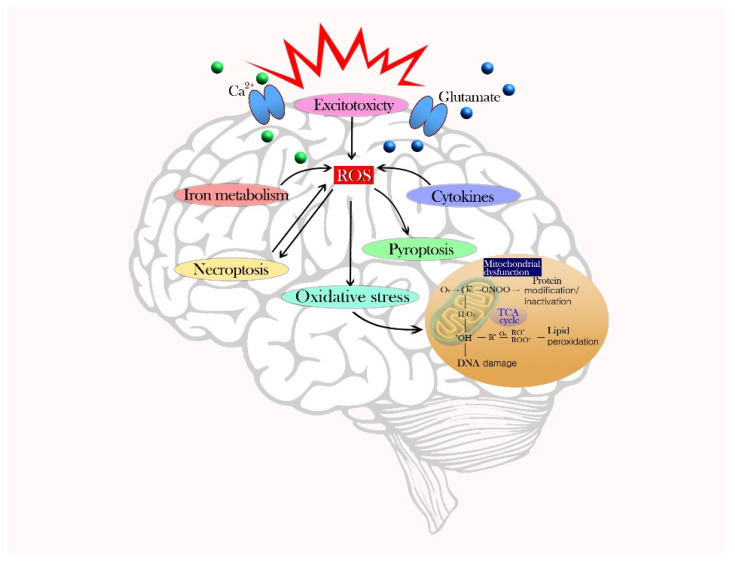
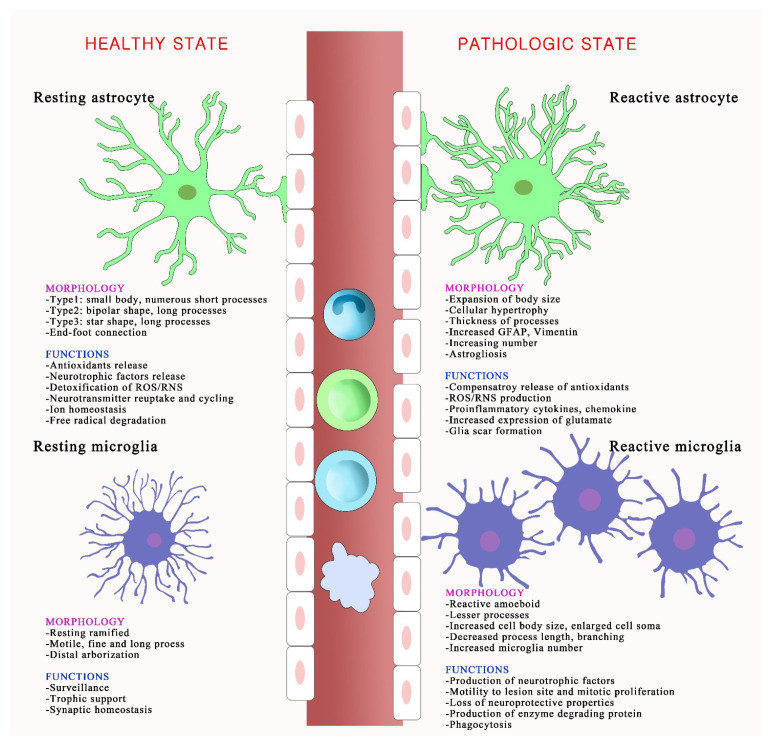
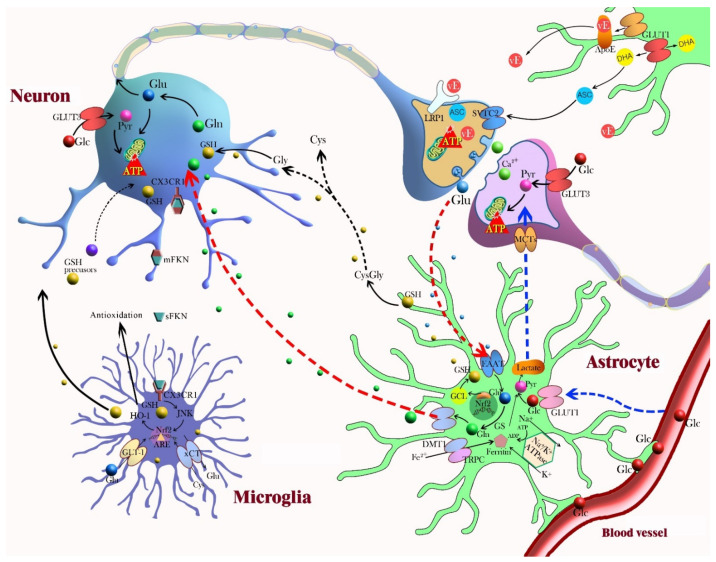
To counteract oxidative stress and associated brain diseases, antioxidant systems rescue neuronal cells from oxidative stress by neutralizing reactive oxygen species and preserving gene regulation. It is necessary to understand the communication and interactions between brain cells, including neurons, astrocytes and microglia, to understand oxidative stress and antioxidant mechanisms. Here, the role of glia in the protection of neurons against oxidative injury and glia-neuron crosstalk to maintain antioxidant defense mechanisms and brain protection are reviewed. The first part of this review focuses on the role of glia in the morphological and physiological changes required for brain homeostasis under oxidative stress and antioxidant defense mechanisms. The second part focuses on the essential crosstalk between neurons and glia for redox balance in the brain for protection against oxidative stress.
Keywords: astrocyte; microglia; neuron–glia interaction; neuroprotection; oxidative injury.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Dual effects of antioxidants in neurodegeneration: direct neuroprotection against oxidative stress and indirect protection via suppression of glia-mediated inflammation.Curr Pharm Des. 2006;12(27):3521-33. doi: 10.2174/138161206778343109. Curr Pharm Des. 2006. PMID: 17017945 Review.
-
Coordinate regulation of glutathione biosynthesis and release by Nrf2-expressing glia potently protects neurons from oxidative stress.J Neurosci. 2003 Apr 15;23(8):3394-406. doi: 10.1523/JNEUROSCI.23-08-03394.2003. J Neurosci. 2003. PMID: 12716947 Free PMC article.
-
Neuron-glia interactions as therapeutic targets in neurodegeneration.J Alzheimers Dis. 2009;16(3):485-502. doi: 10.3233/JAD-2009-0988. J Alzheimers Dis. 2009. PMID: 19276541 Review.
-
Neuron-glia Interaction as a Possible Pathophysiological Mechanism of Bipolar Disorder.Curr Neuropharmacol. 2018;16(5):519-532. doi: 10.2174/1570159X15666170828170921. Curr Neuropharmacol. 2018. PMID: 28847296 Free PMC article. Review.
-
Crosstalk Between the Oxidative Stress and Glia Cells After Stroke: From Mechanism to Therapies.Front Immunol. 2022 Feb 25;13:852416. doi: 10.3389/fimmu.2022.852416. eCollection 2022. Front Immunol. 2022. PMID: 35281064 Free PMC article. Review.
Cited by
-
Lactide and Ethylene Brassylate-Based Thermoplastic Elastomers and Their Nanocomposites with Carbon Nanotubes: Synthesis, Mechanical Properties and Interaction with Astrocytes.Polymers (Basel). 2022 Nov 1;14(21):4656. doi: 10.3390/polym14214656. Polymers (Basel). 2022. PMID: 36365648 Free PMC article.
-
TNF-α Levels Are Increased in Patients with Subjective Cognitive Impairment and Are Negatively Correlated with β Amyloid-42.Antioxidants (Basel). 2024 Feb 8;13(2):216. doi: 10.3390/antiox13020216. Antioxidants (Basel). 2024. PMID: 38397814 Free PMC article.
-
Differential Mitochondrial Bioenergetics in Neurons and Astrocytes Following Ischemia-Reperfusion Injury and Hypothermia.Biomedicines. 2024 Aug 1;12(8):1705. doi: 10.3390/biomedicines12081705. Biomedicines. 2024. PMID: 39200170 Free PMC article.
-
Iron Brain Menace: The Involvement of Ferroptosis in Parkinson Disease.Cells. 2022 Nov 29;11(23):3829. doi: 10.3390/cells11233829. Cells. 2022. PMID: 36497089 Free PMC article. Review.
-
Alzheimer's disease and its treatment-yesterday, today, and tomorrow.Front Pharmacol. 2024 May 24;15:1399121. doi: 10.3389/fphar.2024.1399121. eCollection 2024. Front Pharmacol. 2024. PMID: 38868666 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
