

ADCC-mediating non-neutralizing antibodies can exert immune pressure in early HIV-1 infection
- PMID: 34788337
- PMCID: PMC8598021
- DOI: 10.1371/journal.ppat.1010046
ADCC-mediating non-neutralizing antibodies can exert immune pressure in early HIV-1 infection
Abstract
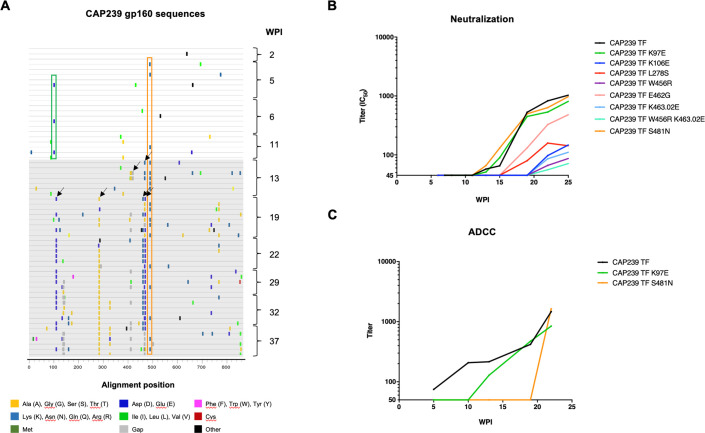
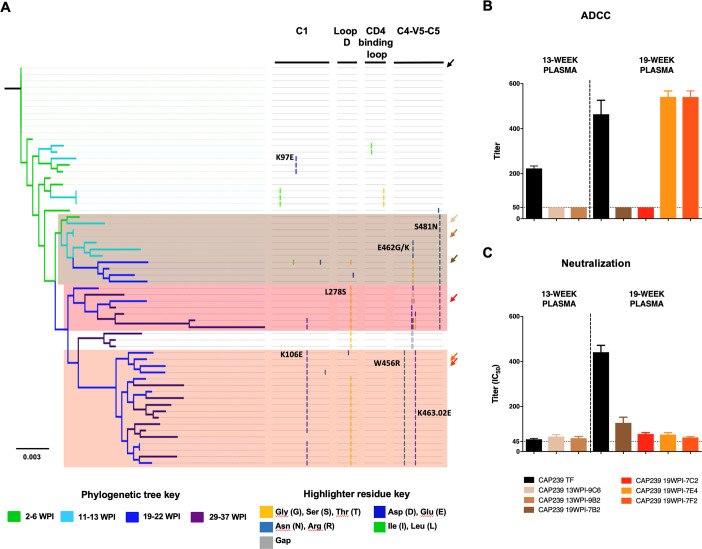
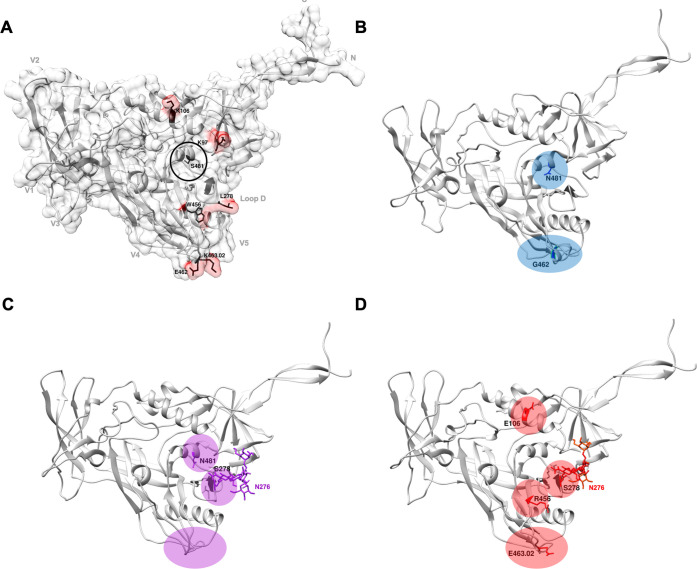
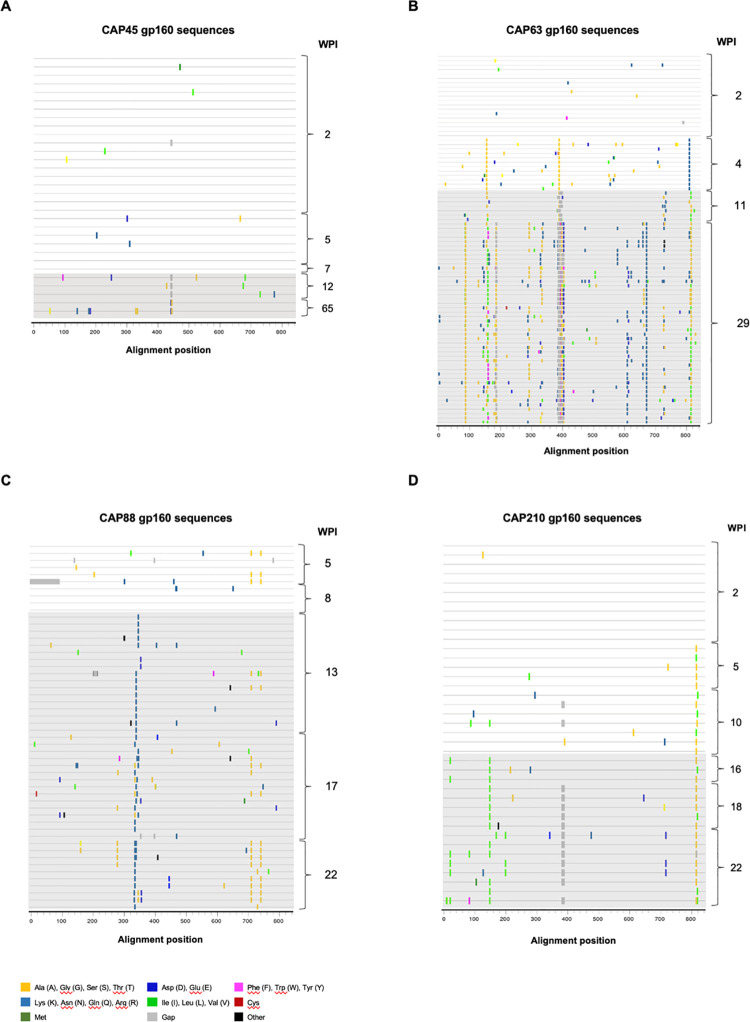
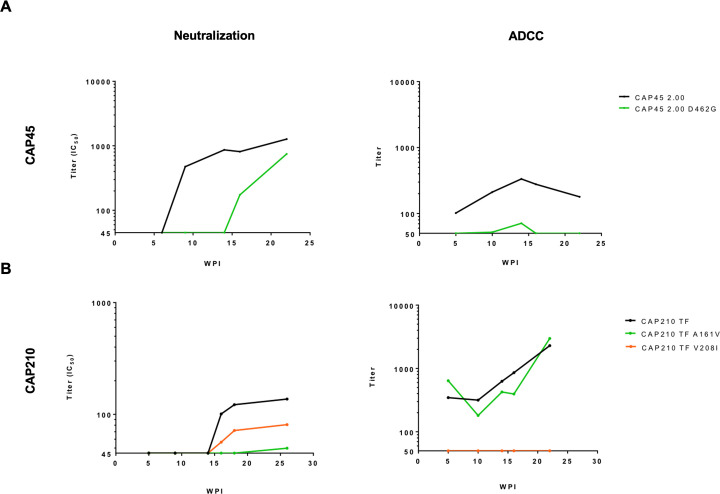
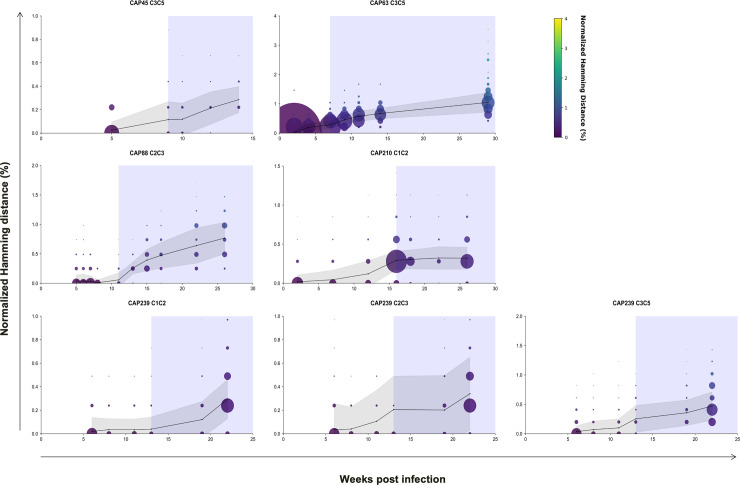
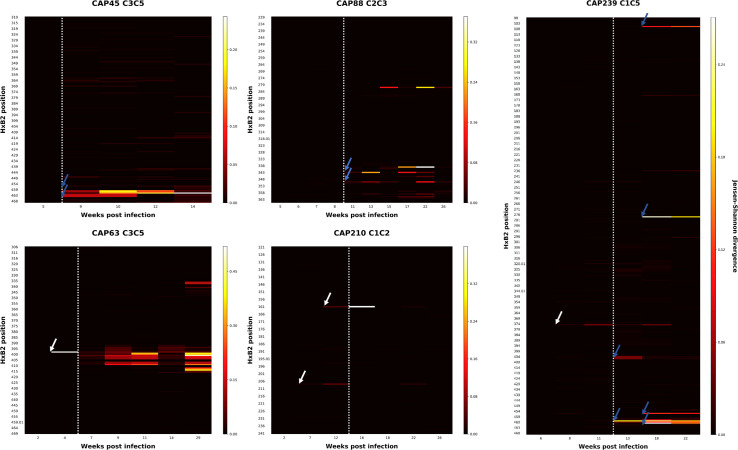
Despite antibody-dependent cellular cytotoxicity (ADCC) responses being implicated in protection from HIV-1 infection, there is limited evidence that they control virus replication. The high mutability of HIV-1 enables the virus to rapidly adapt, and thus evidence of viral escape is a very sensitive approach to demonstrate the importance of this response. To enable us to deconvolute ADCC escape from neutralizing antibody (nAb) escape, we identified individuals soon after infection with detectable ADCC responses, but no nAb responses. We evaluated the kinetics of ADCC and nAb responses, and viral escape, in five recently HIV-1-infected individuals. In one individual we detected viruses that escaped from ADCC responses but were sensitive to nAbs. In the remaining four participants, we did not find evidence of viral evolution exclusively associated with ADCC-mediating non-neutralizing Abs (nnAbs). However, in all individuals escape from nAbs was rapid, occurred at very low titers, and in three of five cases we found evidence of viral escape before detectable nAb responses. These data show that ADCC-mediating nnAbs can drive immune escape in early infection, but that nAbs were far more effective. This suggests that if ADCC responses have a protective role, their impact is limited after systemic virus dissemination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
CD4 downregulation precedes Env expression and protects HIV-1-infected cells from ADCC mediated by non-neutralizing antibodies.mBio. 2024 Nov 13;15(11):e0182724. doi: 10.1128/mbio.01827-24. Epub 2024 Oct 7. mBio. 2024. PMID: 39373535 Free PMC article.
-
Antibody-Dependent Cellular Cytotoxicity (ADCC)-Mediating Antibodies Constrain Neutralizing Antibody Escape Pathway.Front Immunol. 2019 Dec 11;10:2875. doi: 10.3389/fimmu.2019.02875. eCollection 2019. Front Immunol. 2019. PMID: 31921139 Free PMC article.
-
Antibody-Dependent Cellular Cytotoxicity-Competent Antibodies against HIV-1-Infected Cells in Plasma from HIV-Infected Subjects.mBio. 2019 Dec 17;10(6):e02690-19. doi: 10.1128/mBio.02690-19. mBio. 2019. PMID: 31848282 Free PMC article.
-
Anti-HIV-1 antibody-dependent cellular cytotoxicity: is there more to antibodies than neutralization?Curr Opin HIV AIDS. 2018 Mar;13(2):160-166. doi: 10.1097/COH.0000000000000439. Curr Opin HIV AIDS. 2018. PMID: 29194123 Review.
-
Beyond Viral Neutralization.AIDS Res Hum Retroviruses. 2017 Aug;33(8):760-764. doi: 10.1089/AID.2016.0299. Epub 2017 Feb 16. AIDS Res Hum Retroviruses. 2017. PMID: 28084796 Free PMC article. Review.
Cited by
-
A viral vaccine design harnessing prior BCG immunization confers protection against Ebola virus.Front Immunol. 2024 Jul 16;15:1429909. doi: 10.3389/fimmu.2024.1429909. eCollection 2024. Front Immunol. 2024. PMID: 39081315 Free PMC article.
-
The HIV Env Glycoprotein Conformational States on Cells and Viruses.mBio. 2022 Apr 26;13(2):e0182521. doi: 10.1128/mbio.01825-21. Epub 2022 Mar 24. mBio. 2022. PMID: 35323042 Free PMC article. Review.
-
Antibody-dependent cellular cytotoxicity responses and susceptibility influence HIV-1 mother-to-child transmission.JCI Insight. 2022 May 9;7(9):e159435. doi: 10.1172/jci.insight.159435. JCI Insight. 2022. PMID: 35324477 Free PMC article.
-
Highly Sensitive Analysis of Cervical Mucosal HIV-1 Infection Using Reporter Viruses Expressing Secreted Nanoluciferase.Methods Mol Biol. 2024;2807:299-323. doi: 10.1007/978-1-0716-3862-0_21. Methods Mol Biol. 2024. PMID: 38743237
-
CD4 downregulation precedes Env expression and protects HIV-1-infected cells from ADCC mediated by non-neutralizing antibodies.mBio. 2024 Nov 13;15(11):e0182724. doi: 10.1128/mbio.01827-24. Epub 2024 Oct 7. mBio. 2024. PMID: 39373535 Free PMC article.
References
-
- Baum LL, Cassutt KJ, Knigge K, Khattri R, Margolick J, Rinaldo C, et al.. HIV-1 gp120-specific antibody-dependent cell-mediated cytotoxicity correlates with rate of disease progression. J Immunol. 1996;157: 2168–2173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
