

Patterning of vertebrate cardiac progenitor fields by retinoic acid signaling
- PMID: 34665508
- PMCID: PMC8599659
- DOI: 10.1002/dvg.23458
Patterning of vertebrate cardiac progenitor fields by retinoic acid signaling
Abstract
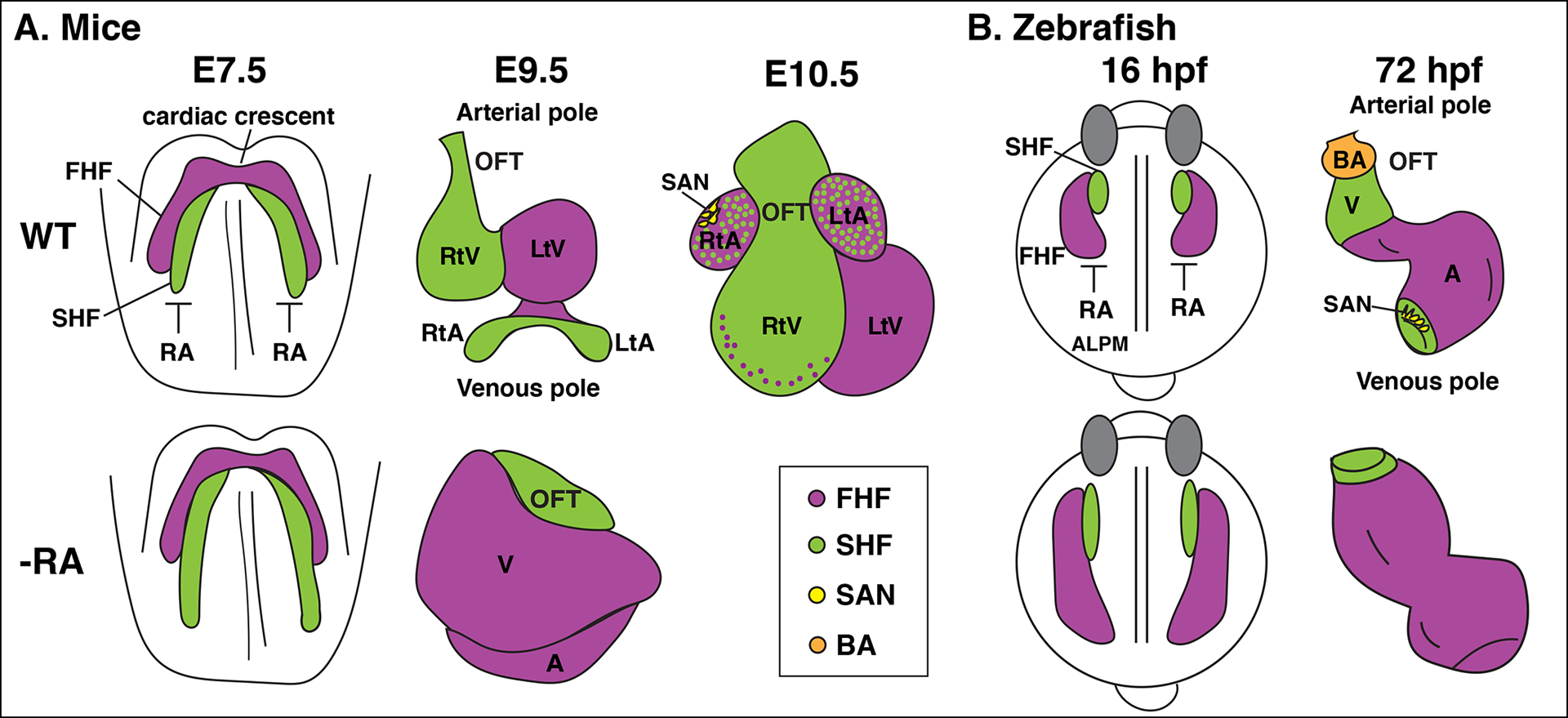
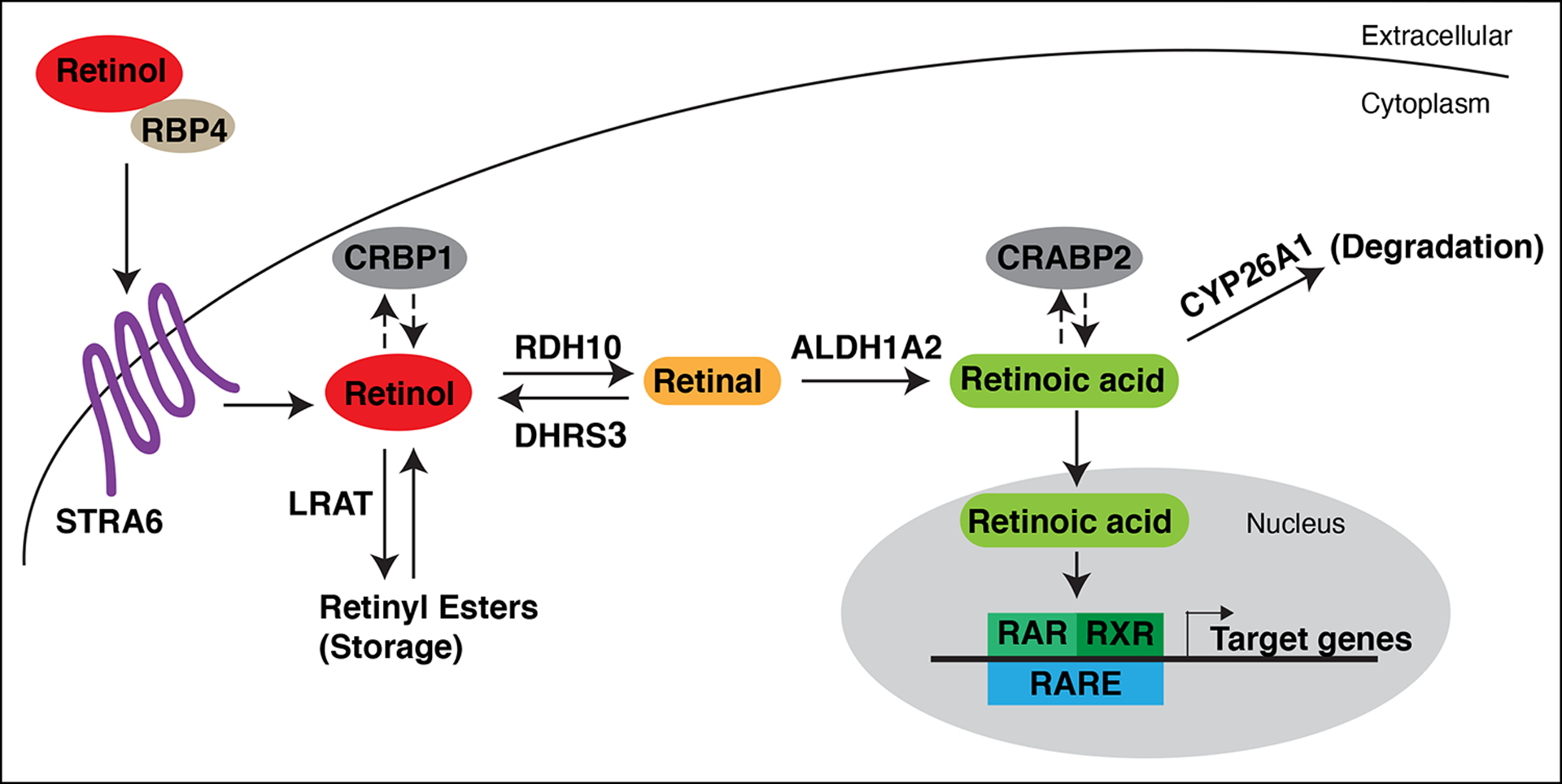
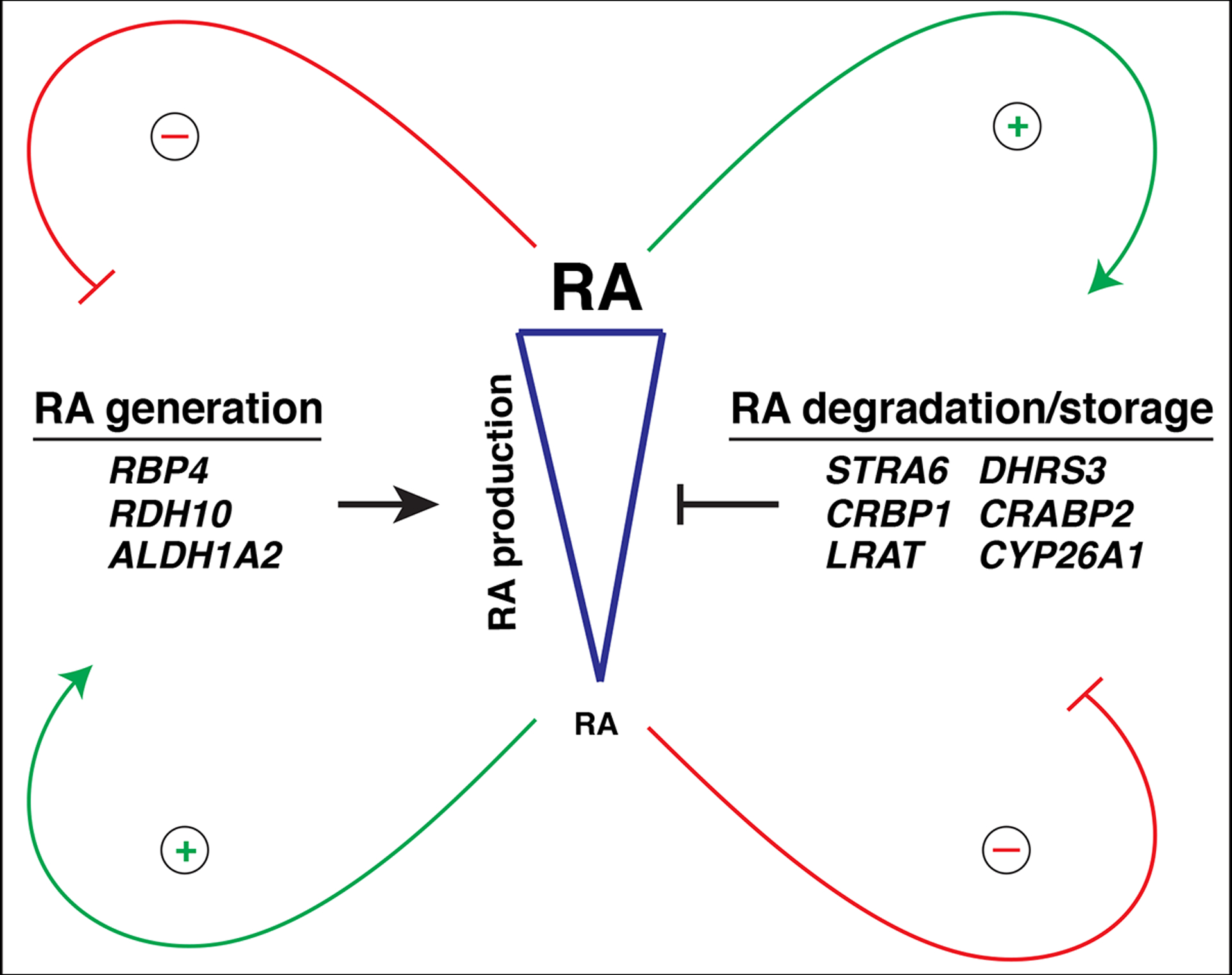
The influence of retinoic acid (RA) signaling on vertebrate development has a well-studied history. Cumulatively, we now understand that RA signaling has a conserved requirement early in development restricting cardiac progenitors within the anterior lateral plate mesoderm of vertebrate embryos. Moreover, genetic and pharmacological manipulations of RA signaling in vertebrate models have shown that proper heart development is achieved through the deployment of positive and negative feedback mechanisms, which maintain appropriate RA levels. In this brief review, we present a chronological overview of key work that has led to a current model of the critical role for early RA signaling in limiting the generation of cardiac progenitors within vertebrate embryos. Furthermore, we integrate the previous work in mice and our recent findings using zebrafish, which together show that RA signaling has remarkably conserved influences on the later-differentiating progenitor populations at the arterial and venous poles. We discuss how recognizing the significant conservation of RA signaling on the differentiation of these progenitor populations offers new perspectives and may impact future work dedicated to examining vertebrate heart development.
Keywords: cardiac progenitors; first heart field; patterning; retinoic acid signaling; second heart field; vertebrate heart development.
© 2021 Wiley Periodicals LLC.
Conflict of interest statement
Figures
Similar articles
-
Retinoic acid signaling restricts the size of the first heart field within the anterior lateral plate mesoderm.Dev Biol. 2021 May;473:119-129. doi: 10.1016/j.ydbio.2021.02.005. Epub 2021 Feb 16. Dev Biol. 2021. PMID: 33607112 Free PMC article.
-
Input overload: Contributions of retinoic acid signaling feedback mechanisms to heart development and teratogenesis.Dev Dyn. 2015 Mar;244(3):513-23. doi: 10.1002/dvdy.24232. Epub 2015 Jan 5. Dev Dyn. 2015. PMID: 25418431 Free PMC article. Review.
-
Retinoic Acid Signaling and Heart Development.Subcell Biochem. 2020;95:119-149. doi: 10.1007/978-3-030-42282-0_5. Subcell Biochem. 2020. PMID: 32297298 Review.
-
T-box genes and retinoic acid signaling regulate the segregation of arterial and venous pole progenitor cells in the murine second heart field.Hum Mol Genet. 2018 Nov 1;27(21):3747-3760. doi: 10.1093/hmg/ddy266. Hum Mol Genet. 2018. PMID: 30016433
-
Restraint of Fgf8 signaling by retinoic acid signaling is required for proper heart and forelimb formation.Dev Biol. 2011 Oct 1;358(1):44-55. doi: 10.1016/j.ydbio.2011.07.022. Epub 2011 Jul 22. Dev Biol. 2011. PMID: 21803036 Free PMC article.
Cited by
-
Inhibition of RhoA and Cdc42 by miR-133a Modulates Retinoic Acid Signalling during Early Development of Posterior Cardiac Tube Segment.Int J Mol Sci. 2022 Apr 10;23(8):4179. doi: 10.3390/ijms23084179. Int J Mol Sci. 2022. PMID: 35456995 Free PMC article.
References
-
- Abu-Abed SS, Beckett BR, Chiba H, Chithalen JV, Jones G, Metzger D, … Petkovich M (1998). Mouse P450RAI (CYP26) expression and retinoic acid-inducible retinoic acid metabolism in F9 cells are regulated by retinoic acid receptor gamma and retinoid X receptor alpha. J Biol Chem, 273(4), 2409–2415. doi:10.1074/jbc.273.4.2409 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
