

Trained innate immunity, long-lasting epigenetic modulation, and skewed myelopoiesis by heme
- PMID: 34663697
- PMCID: PMC8545490
- DOI: 10.1073/pnas.2102698118
Trained innate immunity, long-lasting epigenetic modulation, and skewed myelopoiesis by heme
Abstract
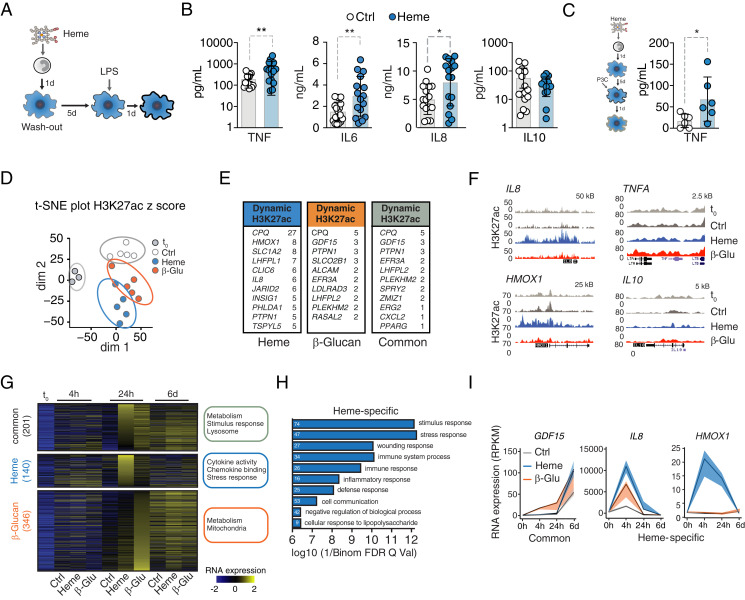
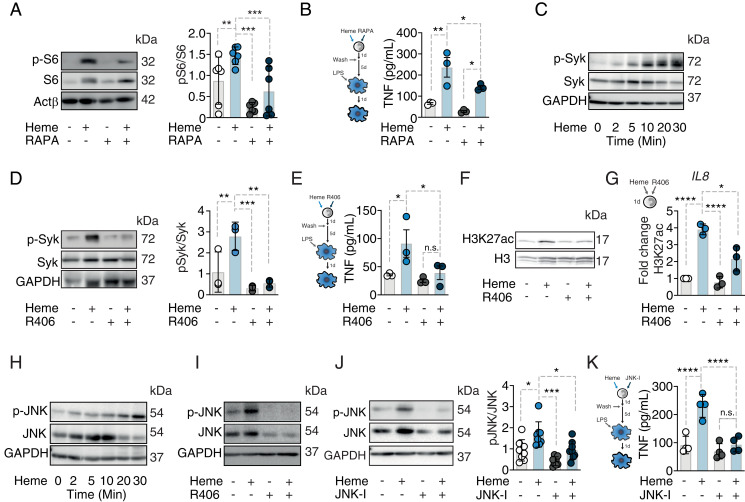
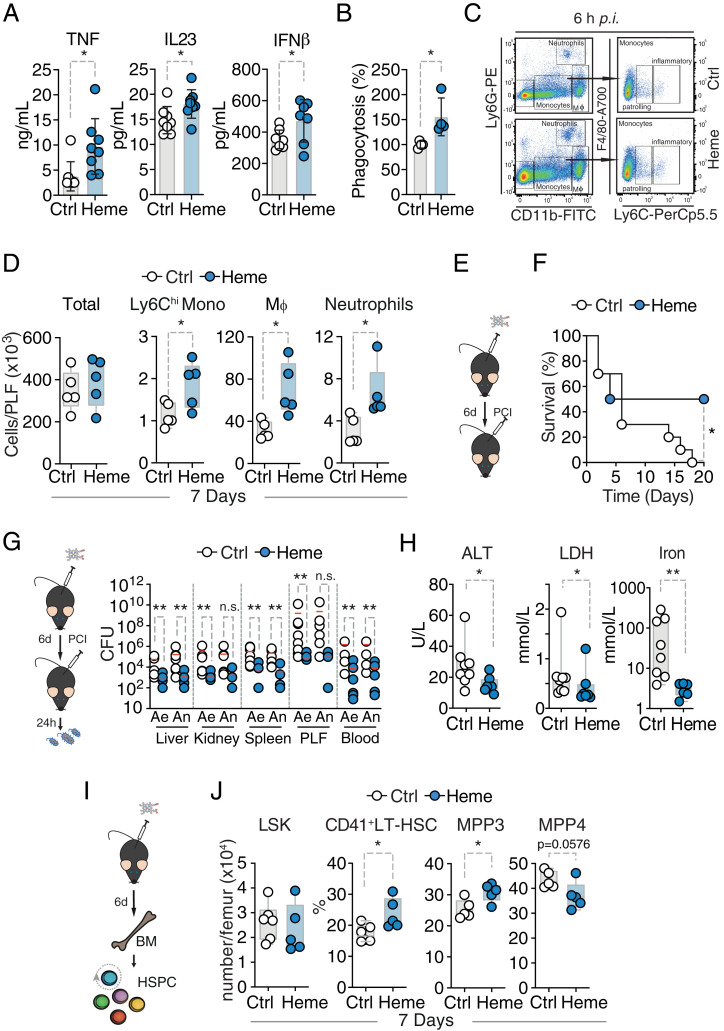
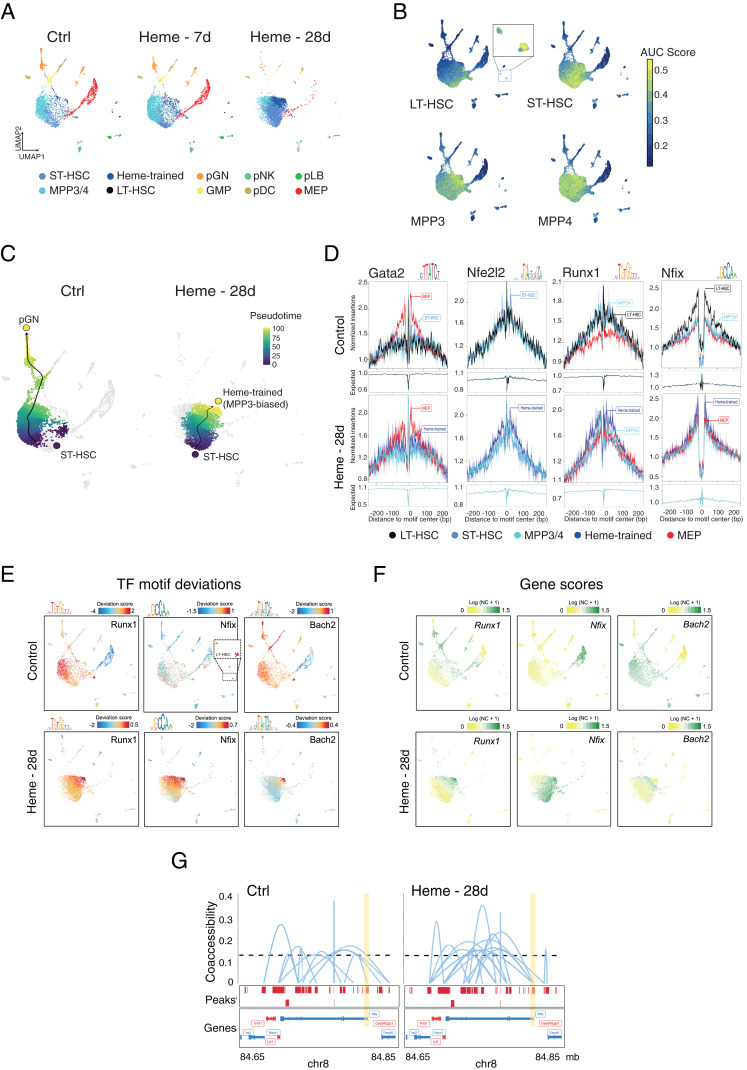
Trained immunity defines long-lasting adaptations of innate immunity based on transcriptional and epigenetic modifications of myeloid cells and their bone marrow progenitors [M. Divangahi et al., Nat. Immunol. 22, 2-6 (2021)]. Innate immune cells, however, do not exclusively differentiate between foreign and self but also react to host-derived molecules referred to as alarmins. Extracellular "labile" heme, released during infections, is a bona fide alarmin promoting myeloid cell activation [M. P. Soares, M. T. Bozza, Curr. Opin. Immunol. 38, 94-100 (2016)]. Here, we report that labile heme is a previously unrecognized inducer of trained immunity that confers long-term regulation of lineage specification of hematopoietic stem cells and progenitor cells. In contrast to previous reports on trained immunity, essentially mediated by pathogen-associated molecular patterns, heme training depends on spleen tyrosine kinase signal transduction pathway acting upstream of c-Jun N-terminal kinases. Heme training promotes resistance to sepsis, is associated with the expansion of self-renewing hematopoetic stem cells primed toward myelopoiesis and to the occurrence of a specific myeloid cell population. This is potentially evoked by sustained activity of Nfix, Runx1, and Nfe2l2 and dissociation of the transcriptional repressor Bach2. Previously reported trained immunity inducers are, however, infrequently present in the host, whereas heme abundantly occurs during noninfectious and infectious disease. This difference might explain the vanishing protection exerted by heme training in sepsis over time with sustained long-term myeloid adaptations. Hence, we propose that trained immunity is an integral component of innate immunity with distinct functional differences on infectious disease outcome depending on its induction by pathogenic or endogenous molecules.
Keywords: heme; myelopoiesis; sepsis; single-nuclei analysis; trained innate immunity.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Modulation of Myelopoiesis Progenitors Is an Integral Component of Trained Immunity.Cell. 2018 Jan 11;172(1-2):147-161.e12. doi: 10.1016/j.cell.2017.11.034. Cell. 2018. PMID: 29328910 Free PMC article.
-
Trained Immunity and Cardiometabolic Disease: The Role of Bone Marrow.Arterioscler Thromb Vasc Biol. 2021 Jan;41(1):48-54. doi: 10.1161/ATVBAHA.120.314215. Epub 2020 Nov 19. Arterioscler Thromb Vasc Biol. 2021. PMID: 33207931 Free PMC article. Review.
-
DAMPs and Innate Immune Training.Front Immunol. 2021 Oct 22;12:699563. doi: 10.3389/fimmu.2021.699563. eCollection 2021. Front Immunol. 2021. PMID: 34745089 Free PMC article. Review.
-
Maladaptive innate immune training of myelopoiesis links inflammatory comorbidities.Cell. 2022 May 12;185(10):1709-1727.e18. doi: 10.1016/j.cell.2022.03.043. Epub 2022 Apr 27. Cell. 2022. PMID: 35483374 Free PMC article.
-
C/EBPβ-Dependent Epigenetic Memory Induces Trained Immunity in Hematopoietic Stem Cells.Cell Stem Cell. 2020 May 7;26(5):657-674.e8. doi: 10.1016/j.stem.2020.01.017. Epub 2020 Mar 12. Cell Stem Cell. 2020. PMID: 32169166
Cited by
-
Multi-Omics Analysis Revealed the rSNPs Potentially Involved in T2DM Pathogenic Mechanism and Metformin Response.Int J Mol Sci. 2024 Aug 27;25(17):9297. doi: 10.3390/ijms25179297. Int J Mol Sci. 2024. PMID: 39273245 Free PMC article.
-
Trained immunity of intestinal tuft cells during infancy enhances host defense against enteroviral infections in mice.EMBO Mol Med. 2024 Oct;16(10):2516-2538. doi: 10.1038/s44321-024-00128-9. Epub 2024 Sep 11. EMBO Mol Med. 2024. PMID: 39261649 Free PMC article.
-
The role of trained immunity in sepsis.Front Immunol. 2024 Aug 15;15:1449986. doi: 10.3389/fimmu.2024.1449986. eCollection 2024. Front Immunol. 2024. PMID: 39221248 Free PMC article. Review.
-
Metabolic Regulation in the Induction of Trained Immunity.Semin Immunopathol. 2024 Jul 25;46(3-4):7. doi: 10.1007/s00281-024-01015-8. Semin Immunopathol. 2024. PMID: 39060761 Free PMC article. Review.
-
The causes and consequences of trained immunity in myeloid cells.Front Immunol. 2024 Apr 11;15:1365127. doi: 10.3389/fimmu.2024.1365127. eCollection 2024. Front Immunol. 2024. PMID: 38665915 Free PMC article. Review.
References
-
- Englert F. A., et al. ., Labile heme impairs hepatic microcirculation and promotes hepatic injury. Arch. Biochem. Biophys. 672, 108075 (2019). - PubMed
-
- Ferreira A., Balla J., Jeney V., Balla G., Soares M. P., A central role for free heme in the pathogenesis of severe malaria: The missing link? J. Mol. Med. (Berl.) 86, 1097–1111 (2008). - PubMed
-
- Larsen R., et al. ., A central role for free heme in the pathogenesis of severe sepsis. Sci. Transl. Med. 2, 51ra71 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
