

mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern
- PMID: 34648302
- PMCID: PMC9284784
- DOI: 10.1126/science.abm0829
mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern
Abstract
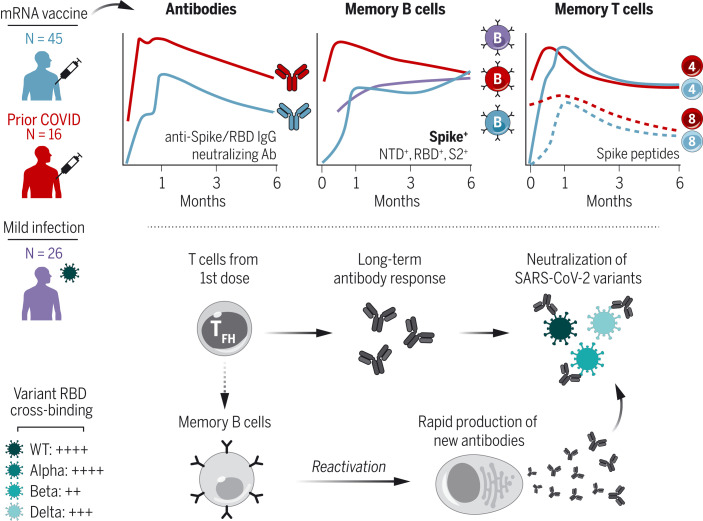
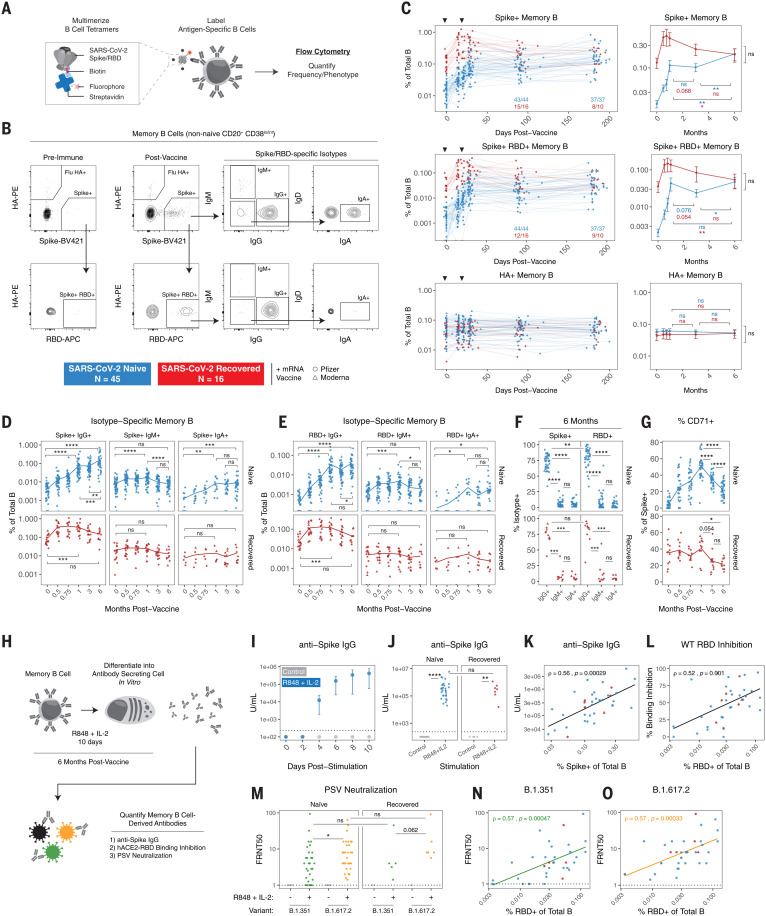
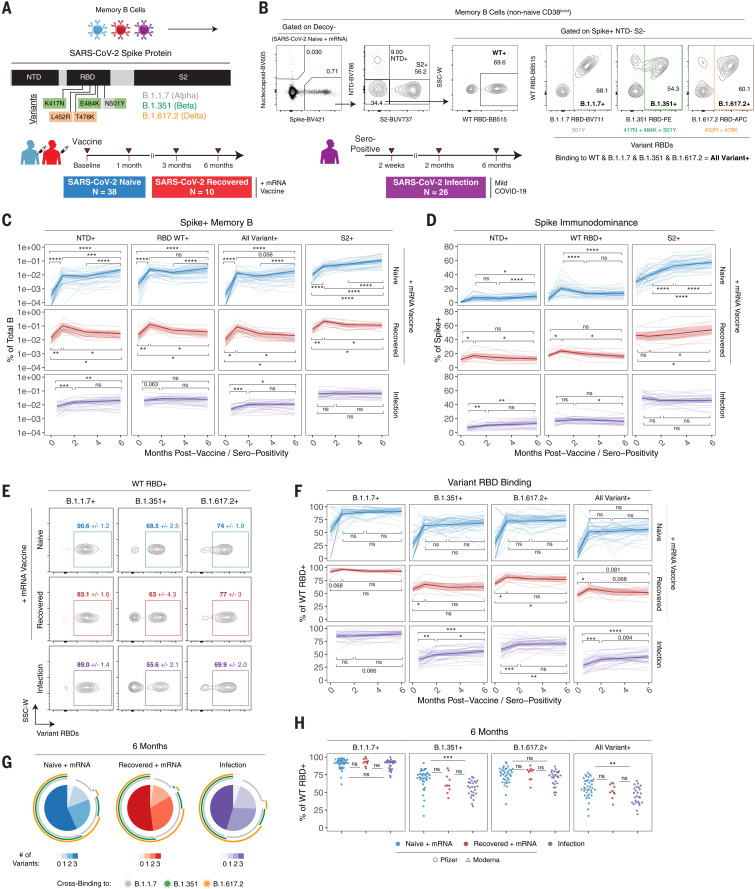
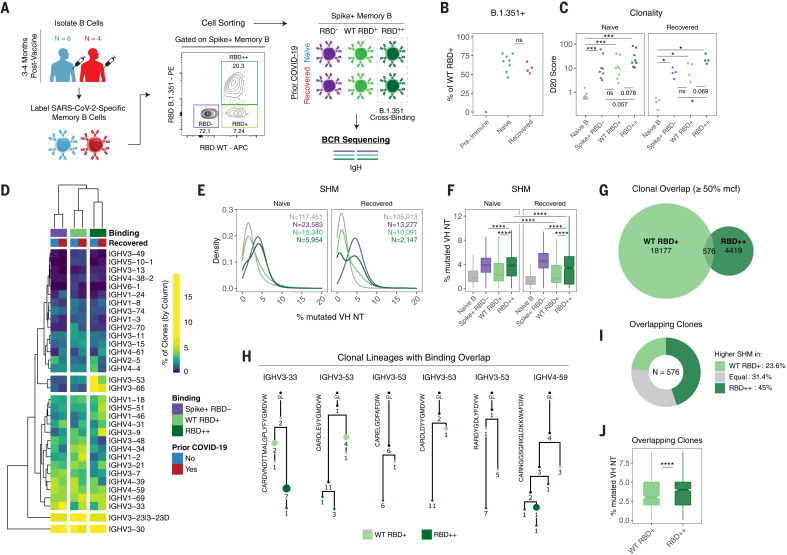
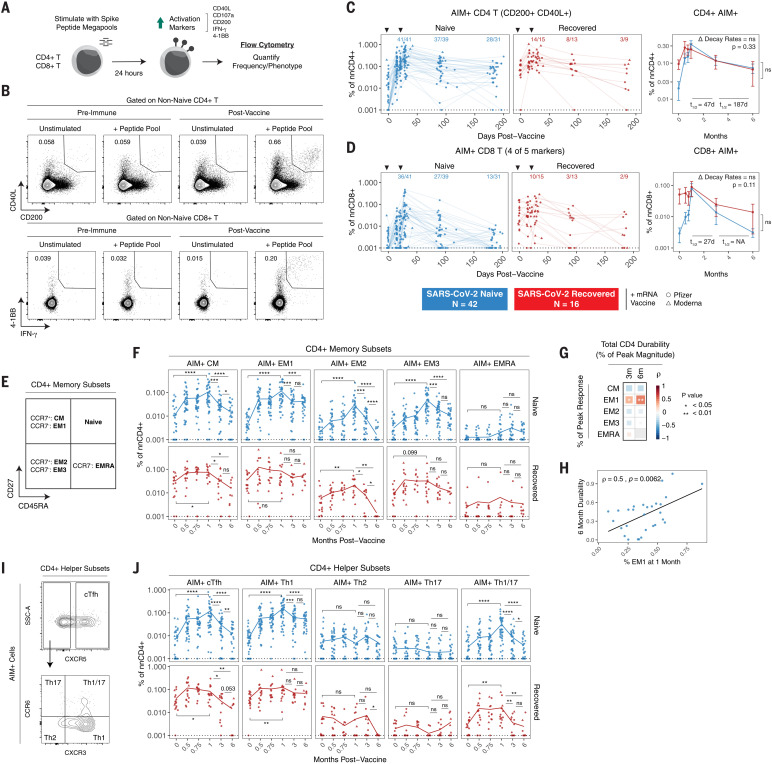
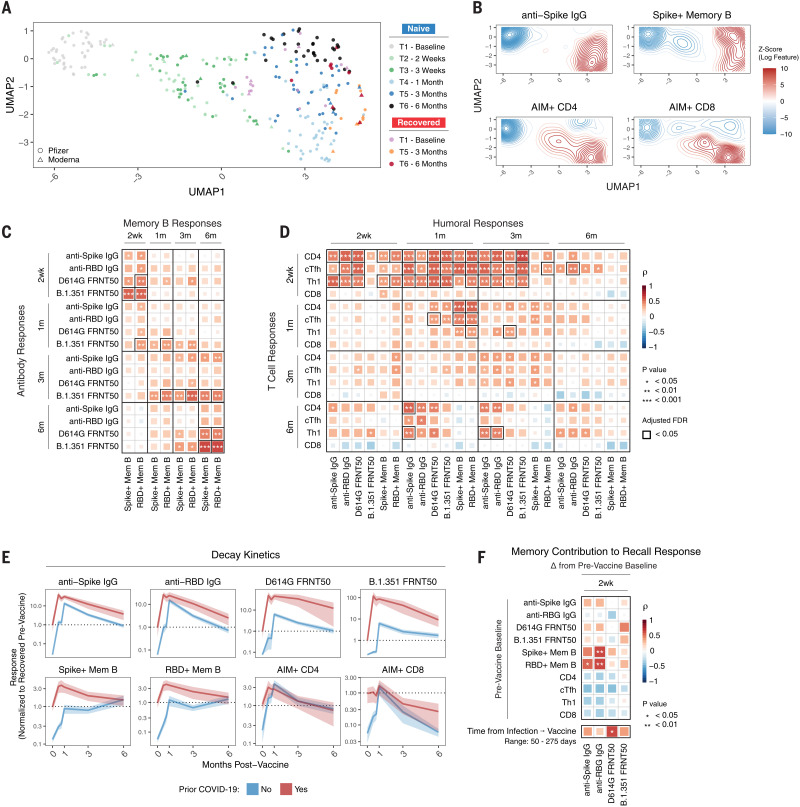
The durability of immune memory after severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) messenger RNA (mRNA) vaccination remains unclear. In this study, we longitudinally profiled vaccine responses in SARS-CoV-2–naïve and –recovered individuals for 6 months after vaccination. Antibodies declined from peak levels but remained detectable in most subjects at 6 months. By contrast, mRNA vaccines generated functional memory B cells that increased from 3 to 6 months postvaccination, with the majority of these cells cross-binding the Alpha, Beta, and Delta variants. mRNA vaccination further induced antigen-specific CD4+ and CD8+ T cells, and early CD4+ T cell responses correlated with long-term humoral immunity. Recall responses to vaccination in individuals with preexisting immunity primarily increased antibody levels without substantially altering antibody decay rates. Together, these findings demonstrate robust cellular immune memory to SARS-CoV-2 and its variants for at least 6 months after mRNA vaccination.
Figures

Update of
-
mRNA Vaccination Induces Durable Immune Memory to SARS-CoV-2 with Continued Evolution to Variants of Concern.bioRxiv [Preprint]. 2021 Aug 23:2021.08.23.457229. doi: 10.1101/2021.08.23.457229. bioRxiv. 2021. Update in: Science. 2021 Dec 03;374(6572):abm0829. doi: 10.1126/science.abm0829 PMID: 34462751 Free PMC article. Updated. Preprint.
Comment in
-
The benefit of boosters: diversity and inclusion in the COVID-19 memory response.Immunol Cell Biol. 2022 Jan;100(1):15-17. doi: 10.1111/imcb.12511. Epub 2021 Dec 5. Immunol Cell Biol. 2022. PMID: 34865234
Similar articles
-
mRNA Vaccination Induces Durable Immune Memory to SARS-CoV-2 with Continued Evolution to Variants of Concern.bioRxiv [Preprint]. 2021 Aug 23:2021.08.23.457229. doi: 10.1101/2021.08.23.457229. bioRxiv. 2021. Update in: Science. 2021 Dec 03;374(6572):abm0829. doi: 10.1126/science.abm0829 PMID: 34462751 Free PMC article. Updated. Preprint.
-
Cellular and humoral functional responses after BNT162b2 mRNA vaccination differ longitudinally between naive and subjects recovered from COVID-19.Cell Rep. 2022 Jan 11;38(2):110235. doi: 10.1016/j.celrep.2021.110235. Epub 2021 Dec 21. Cell Rep. 2022. PMID: 34986327 Free PMC article.
-
SARS-CoV-2 mRNA vaccine induces robust specific and cross-reactive IgG and unequal neutralizing antibodies in naive and previously infected people.Cell Rep. 2022 Feb 1;38(5):110336. doi: 10.1016/j.celrep.2022.110336. Epub 2022 Jan 20. Cell Rep. 2022. PMID: 35090596 Free PMC article.
-
SARS-CoV-2 Variants, Vaccines, and Host Immunity.Front Immunol. 2022 Jan 3;12:809244. doi: 10.3389/fimmu.2021.809244. eCollection 2021. Front Immunol. 2022. PMID: 35046961 Free PMC article. Review.
-
The germinal centre B cell response to SARS-CoV-2.Nat Rev Immunol. 2022 Jan;22(1):7-18. doi: 10.1038/s41577-021-00657-1. Epub 2021 Dec 6. Nat Rev Immunol. 2022. PMID: 34873279 Free PMC article. Review.
Cited by
-
Early CD4+ T cell responses induced by the BNT162b2 SARS-CoV-2 mRNA vaccine predict immunological memory.Sci Rep. 2022 Nov 27;12(1):20376. doi: 10.1038/s41598-022-24938-4. Sci Rep. 2022. PMID: 36437407 Free PMC article.
-
A systematic and thematic analysis of the top 100 cited articles on mRNA vaccine indexed in Scopus database.Hum Vaccin Immunother. 2022 Nov 30;18(6):2135927. doi: 10.1080/21645515.2022.2135927. Epub 2022 Nov 3. Hum Vaccin Immunother. 2022. PMID: 36328513 Free PMC article.
-
Human RP105 monoclonal antibody enhances antigen-specific antibody production in unique culture conditions.iScience. 2024 Aug 3;27(9):110649. doi: 10.1016/j.isci.2024.110649. eCollection 2024 Sep 20. iScience. 2024. PMID: 39246445 Free PMC article.
-
Atypical and non-classical CD45RBlo memory B cells are the majority of circulating SARS-CoV-2 specific B cells following mRNA vaccination or COVID-19.Nat Commun. 2024 Aug 9;15(1):6811. doi: 10.1038/s41467-024-50997-4. Nat Commun. 2024. PMID: 39122676 Free PMC article.
-
Feline Infectious Peritonitis mRNA Vaccine Elicits Both Humoral and Cellular Immune Responses in Mice.Vaccines (Basel). 2024 Jun 24;12(7):705. doi: 10.3390/vaccines12070705. Vaccines (Basel). 2024. PMID: 39066343 Free PMC article.
References
-
- Polack F. P., Thomas S. J., Kitchin N., Absalon J., Gurtman A., Lockhart S., Perez J. L., Pérez Marc G., Moreira E. D., Zerbini C., Bailey R., Swanson K. A., Roychoudhury S., Koury K., Li P., Kalina W. V., Cooper D., Frenck R. W., Hammitt L. L., Türeci Ö., Nell H., Schaefer A., Ünal S., Tresnan D. B., Mather S., Dormitzer P. R., Şahin U., Jansen K. U., Gruber W. C., Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 383, 2603–2615 (2020). 10.1056/NEJMoa2034577 - DOI - PMC - PubMed
-
- Baden L. R., El Sahly H. M., Essink B., Kotloff K., Frey S., Novak R., Diemert D., Spector S. A., Rouphael N., Creech C. B., McGettigan J., Khetan S., Segall N., Solis J., Brosz A., Fierro C., Schwartz H., Neuzil K., Corey L., Gilbert P., Janes H., Follmann D., Marovich M., Mascola J., Polakowski L., Ledgerwood J., Graham B. S., Bennett H., Pajon R., Knightly C., Leav B., Deng W., Zhou H., Han S., Ivarsson M., Miller J., Zaks T., Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 384, 403–416 (2021). 10.1056/NEJMoa2035389 - DOI - PMC - PubMed
-
- Khoury D. S., Cromer D., Reynaldi A., Schlub T. E., Wheatley A. K., Juno J. A., Subbarao K., Kent S. J., Triccas J. A., Davenport M. P., Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 27, 1205–1211 (2021). 10.1038/s41591-021-01377-8 - DOI - PubMed
-
- Cromer D., Steain M., Reynaldi A., Schlub T. E., Wheatley A. K., Juno J. A., Kent S. J., Triccas J. A., Khoury D. S., Davenport M. P., SARS-CoV-2 variants: levels of neutralisation required for protective immunity. medRxiv 2021.08.11.21261876 [Preprint] (2021). .10.1101/2021.08.11.21261876 - DOI
Publication types
MeSH terms
Substances
Grants and funding
- T32 AR076951/AR/NIAMS NIH HHS/United States
- R21 AI142638/AI/NIAID NIH HHS/United States
- R01 AI152236/AI/NIAID NIH HHS/United States
- T32 AI070077/AI/NIAID NIH HHS/United States
- T32 CA009140/CA/NCI NIH HHS/United States
- P30 AI045008/AI/NIAID NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- R38 HL143613/HL/NHLBI NIH HHS/United States
- P01 AI108545/AI/NIAID NIH HHS/United States
- R01 AI105343/AI/NIAID NIH HHS/United States
- T32 AI055400/AI/NIAID NIH HHS/United States
- R01 AI155577/AI/NIAID NIH HHS/United States
- U19 AI149680/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
