

In situ cryo-electron tomography reveals gradient organization of ribosome biogenesis in intact nucleoli
- PMID: 34508074
- PMCID: PMC8433212
- DOI: 10.1038/s41467-021-25413-w
In situ cryo-electron tomography reveals gradient organization of ribosome biogenesis in intact nucleoli
Abstract
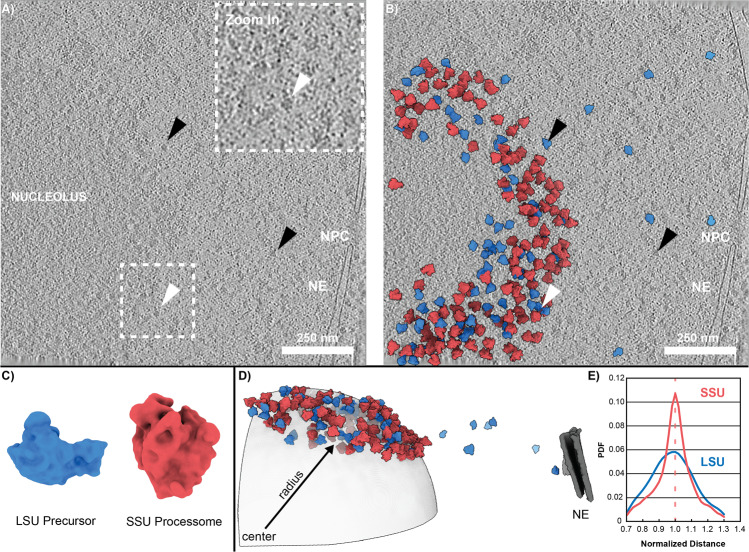
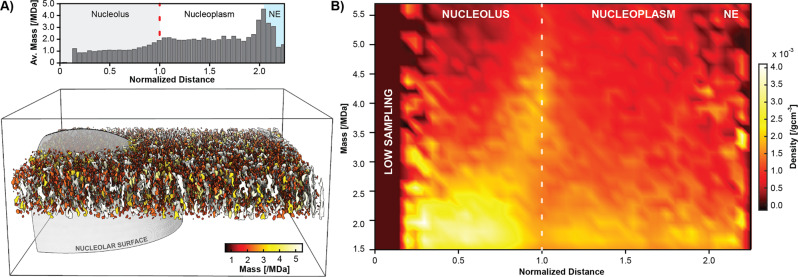
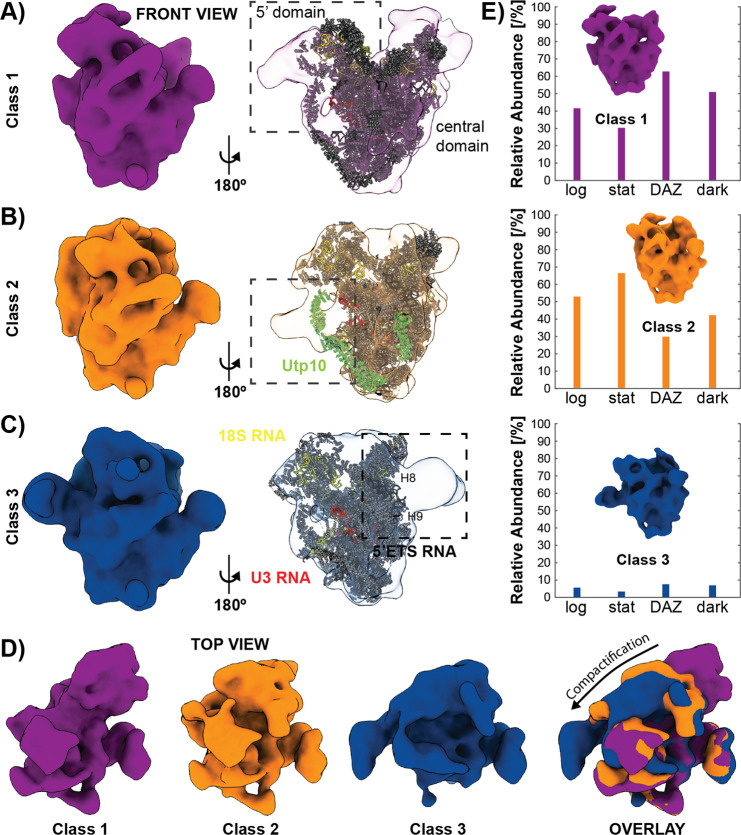
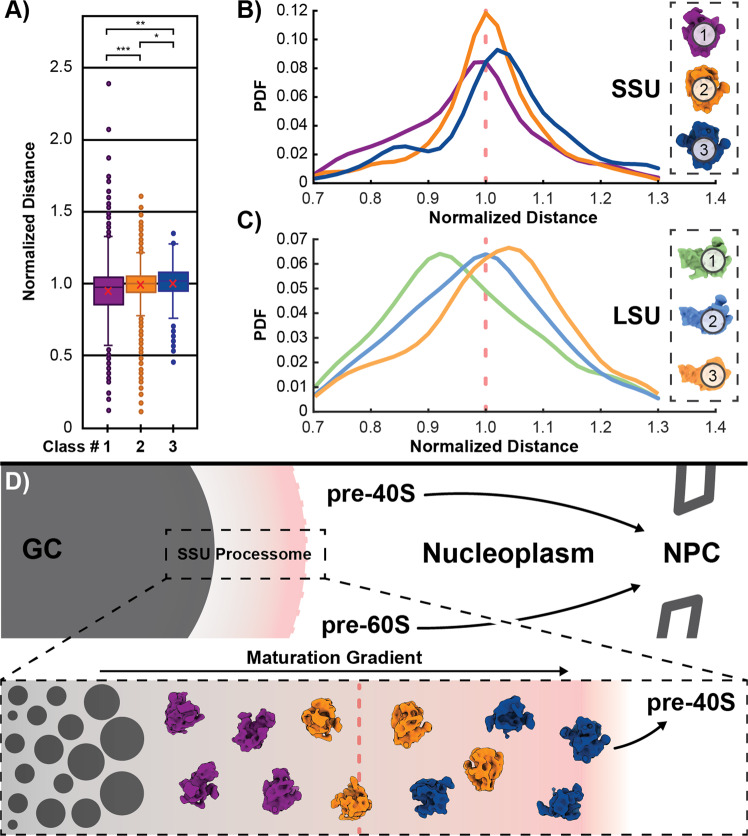
Ribosomes comprise a large (LSU) and a small subunit (SSU) which are synthesized independently in the nucleolus before being exported into the cytoplasm, where they assemble into functional ribosomes. Individual maturation steps have been analyzed in detail using biochemical methods, light microscopy and conventional electron microscopy (EM). In recent years, single particle analysis (SPA) has yielded molecular resolution structures of several pre-ribosomal intermediates. It falls short, however, of revealing the spatiotemporal sequence of ribosome biogenesis in the cellular context. Here, we present our study on native nucleoli in Chlamydomonas reinhardtii, in which we follow the formation of LSU and SSU precursors by in situ cryo-electron tomography (cryo-ET) and subtomogram averaging (STA). By combining both positional and molecular data, we reveal gradients of ribosome maturation within the granular component (GC), offering a new perspective on how the liquid-liquid-phase separation of the nucleolus supports ribosome biogenesis.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Nucleolus: the ribosome factory.Histol Histopathol. 2008 Oct;23(10):1291-8. doi: 10.14670/HH-23.1291. Histol Histopathol. 2008. PMID: 18712681 Review.
-
Eukaryotic Ribosome Assembly.Annu Rev Biochem. 2019 Jun 20;88:281-306. doi: 10.1146/annurev-biochem-013118-110817. Epub 2018 Dec 19. Annu Rev Biochem. 2019. PMID: 30566372 Review.
-
Good Vibrations: Structural Remodeling of Maturing Yeast Pre-40S Ribosomal Particles Followed by Cryo-Electron Microscopy.Molecules. 2020 Mar 3;25(5):1125. doi: 10.3390/molecules25051125. Molecules. 2020. PMID: 32138239 Free PMC article.
-
Intranucleolar sites of ribosome biogenesis defined by the localization of early binding ribosomal proteins.J Cell Biol. 2007 May 21;177(4):573-8. doi: 10.1083/jcb.200612048. J Cell Biol. 2007. PMID: 17517959 Free PMC article.
-
A comprehensive landscape of 60S ribosome biogenesis factors.Cell Rep. 2022 Feb 8;38(6):110353. doi: 10.1016/j.celrep.2022.110353. Cell Rep. 2022. PMID: 35139378 Free PMC article.
Cited by
-
Mapping and engineering RNA-controlled architecture of the multiphase nucleolus.bioRxiv [Preprint]. 2024 Sep 29:2024.09.28.615444. doi: 10.1101/2024.09.28.615444. bioRxiv. 2024. PMID: 39386460 Free PMC article. Preprint.
-
Emergence of the primordial pre-60S from the 90S pre-ribosome.Cell Rep. 2022 Apr 5;39(1):110640. doi: 10.1016/j.celrep.2022.110640. Cell Rep. 2022. PMID: 35385737 Free PMC article.
-
Inherently disordered regions of axonemal dynein assembly factors.Cytoskeleton (Hoboken). 2024 Nov;81(11):515-528. doi: 10.1002/cm.21789. Epub 2023 Sep 15. Cytoskeleton (Hoboken). 2024. PMID: 37712517 Review.
-
Ultrastructural insights into cellular organization, energy storage and ribosomal dynamics of an ammonia-oxidizing archaeon from oligotrophic oceans.Front Microbiol. 2024 Apr 26;15:1367658. doi: 10.3389/fmicb.2024.1367658. eCollection 2024. Front Microbiol. 2024. PMID: 38737410 Free PMC article.
-
A method for restoring signals and revealing individual macromolecule states in cryo-ET, REST.Nat Commun. 2023 May 22;14(1):2937. doi: 10.1038/s41467-023-38539-w. Nat Commun. 2023. PMID: 37217501 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
