

Immunodominant Cytomegalovirus Epitopes Suppress Subdominant Epitopes in the Generation of High-Avidity CD8 T Cells
- PMID: 34451420
- PMCID: PMC8400798
- DOI: 10.3390/pathogens10080956
Immunodominant Cytomegalovirus Epitopes Suppress Subdominant Epitopes in the Generation of High-Avidity CD8 T Cells
Abstract
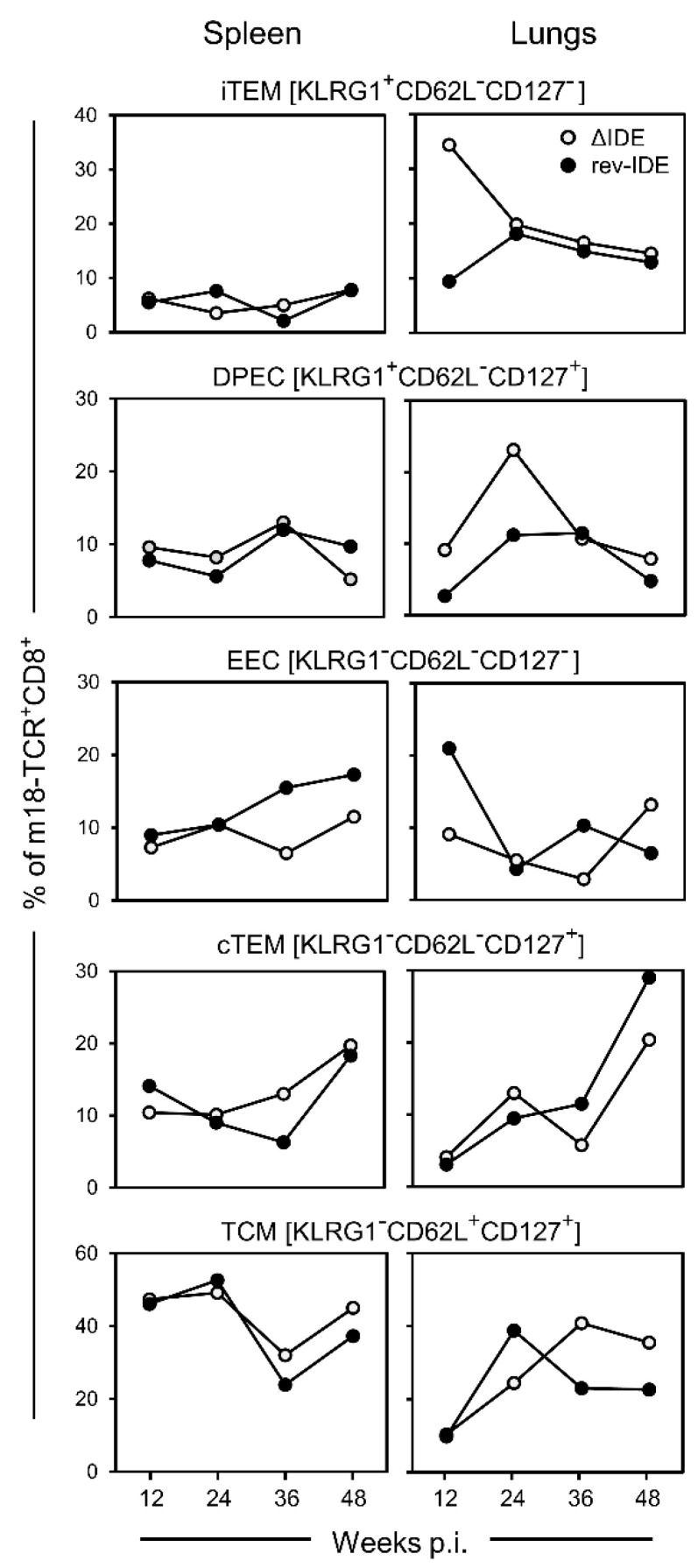
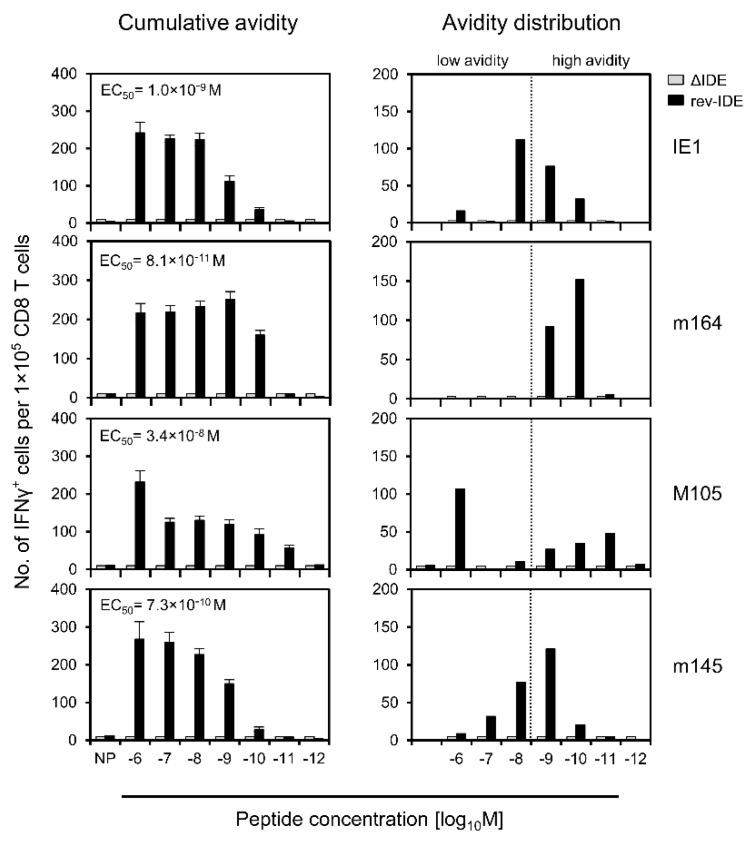
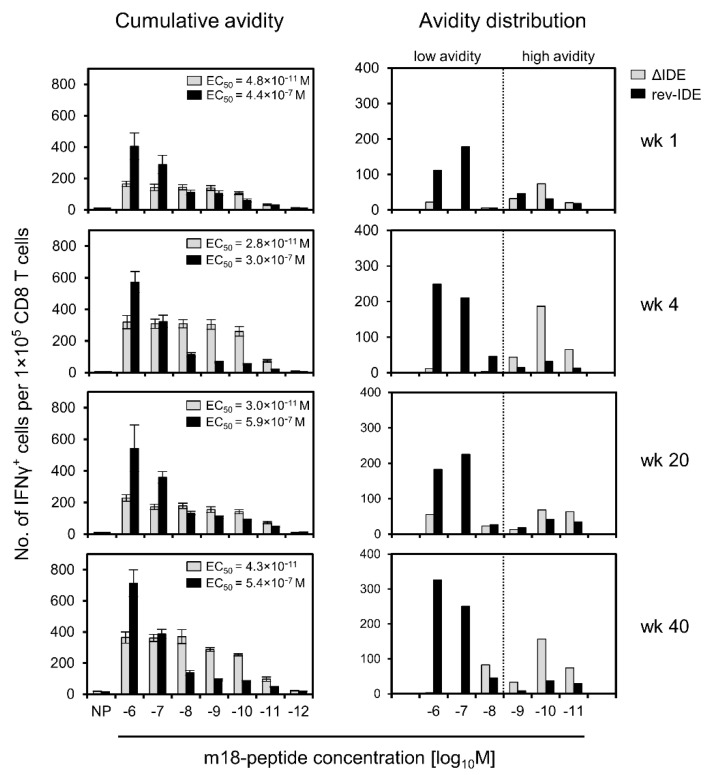
CD8+ T-cell responses to pathogens are directed against infected cells that present pathogen-encoded peptides on MHC class-I molecules. Although natural responses are polyclonal, the spectrum of peptides that qualify for epitopes is remarkably small even for pathogens with high coding capacity. Among those few that are successful at all, a hierarchy exists in the magnitude of the response that they elicit in terms of numbers of CD8+ T cells generated. This led to a classification into immunodominant and non-immunodominant or subordinate epitopes, IDEs and non-IDEs, respectively. IDEs are favored in the design of vaccines and are chosen for CD8+ T-cell immunotherapy. Using murine cytomegalovirus as a model, we provide evidence to conclude that epitope hierarchy reflects competition on the level of antigen recognition. Notably, high-avidity cells specific for non-IDEs were found to expand only when IDEs were deleted. This may be a host's back-up strategy to avoid viral immune escape through antigenic drift caused by IDE mutations. Importantly, our results are relevant for the design of vaccines based on cytomegaloviruses as vectors to generate high-avidity CD8+ T-cell memory specific for unrelated pathogens or tumors. We propose the deletion of vector-encoded IDEs to avoid the suppression of epitopes of the vaccine target.
Keywords: CD8 T cells; antigen presentation; antigenic peptides; cytomegalovirus; epitope(s); immunodominance; immunotherapy; protective immunity; vaccine design.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript or in the decision to publish the results.
Figures


Similar articles
-
Reconstitution of CD8 T Cells Protective against Cytomegalovirus in a Mouse Model of Hematopoietic Cell Transplantation: Dynamics and Inessentiality of Epitope Immunodominance.Front Immunol. 2016 Jun 14;7:232. doi: 10.3389/fimmu.2016.00232. eCollection 2016. Front Immunol. 2016. PMID: 27379095 Free PMC article.
-
Immune control in the absence of immunodominant epitopes: implications for immunotherapy of cytomegalovirus infection with antiviral CD8 T cells.Med Microbiol Immunol. 2012 Nov;201(4):541-50. doi: 10.1007/s00430-012-0268-8. Epub 2012 Sep 14. Med Microbiol Immunol. 2012. PMID: 22976556
-
Subdominant CD8 T-cell epitopes account for protection against cytomegalovirus independent of immunodomination.J Virol. 2008 Jun;82(12):5781-96. doi: 10.1128/JVI.00155-08. Epub 2008 Mar 26. J Virol. 2008. PMID: 18367531 Free PMC article.
-
Protein antigenic structures recognized by T cells: potential applications to vaccine design.Immunol Rev. 1987 Aug;98:9-52. doi: 10.1111/j.1600-065x.1987.tb00518.x. Immunol Rev. 1987. PMID: 2443443 Review.
-
Revealing factors determining immunodominant responses against dominant epitopes.Immunogenetics. 2020 Feb;72(1-2):109-118. doi: 10.1007/s00251-019-01134-9. Epub 2019 Dec 6. Immunogenetics. 2020. PMID: 31811313 Free PMC article. Review.
Cited by
-
Immunotherapy of cytomegalovirus infection by low-dose adoptive transfer of antiviral CD8 T cells relies on substantial post-transfer expansion of central memory cells but not effector-memory cells.PLoS Pathog. 2023 Nov 16;19(11):e1011643. doi: 10.1371/journal.ppat.1011643. eCollection 2023 Nov. PLoS Pathog. 2023. PMID: 37972198 Free PMC article.
-
Modulation of cytomegalovirus immune evasion identifies direct antigen presentation as the predominant mode of CD8 T-cell priming during immune reconstitution after hematopoietic cell transplantation.Front Immunol. 2024 Feb 15;15:1355153. doi: 10.3389/fimmu.2024.1355153. eCollection 2024. Front Immunol. 2024. PMID: 38426094 Free PMC article.
-
A robust deep learning workflow to predict CD8 + T-cell epitopes.Genome Med. 2023 Sep 13;15(1):70. doi: 10.1186/s13073-023-01225-z. Genome Med. 2023. PMID: 37705109 Free PMC article.
References
-
- Davison A.J., Holton M., Dolan A., Dargan D.J., Gatherer D., Hayward G.S. Comparative genomics of primate cytomegaloviruses. In: Reddehase M.J., editor. Cytomegaloviruses: From Molecular Pathogenesis to Intervention. Volume I. Caister Academic Press; Norfolk, UK: 2013. pp. 1–22.
-
- Redwood A.J., Shellam G.R., Smith L.M. Molecular evolution of murine cytomegalovirus genomes. In: Reddehase M.J., editor. Cytomegaloviruses: From Molecular Pathogenesis to Intervention. Volume I. Caister Academic Press; Norfolk, UK: 2013. pp. 23–37.
-
- Cannon M.J., Grosse S.D., Fowler K.B. The epidemiology and public health impact of congenital cytomegalovirus infection. In: Reddehase M.J., editor. Cytomegaloviruses: From Molecular Pathogenesis to Intervention. Volume II. Caister Academic Press; Norfolk, UK: 2013. pp. 26–48.
-
- Adler S.P., Nigro G. Clinical cytomegalovirus research: Congential infection. In: Reddehase M.J., editor. Cytomegaloviruses: From Molecular Pathogenesis to Intervention. Volume II. Caister Academic Press; Norfolk, UK: 2013. pp. 55–73.
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
