

Natural experiments and long-term monitoring are critical to understand and predict marine host-microbe ecology and evolution
- PMID: 34411089
- PMCID: PMC8376202
- DOI: 10.1371/journal.pbio.3001322
Natural experiments and long-term monitoring are critical to understand and predict marine host-microbe ecology and evolution
Abstract
Marine multicellular organisms host a diverse collection of bacteria, archaea, microbial eukaryotes, and viruses that form their microbiome. Such host-associated microbes can significantly influence the host's physiological capacities; however, the identity and functional role(s) of key members of the microbiome ("core microbiome") in most marine hosts coexisting in natural settings remain obscure. Also unclear is how dynamic interactions between hosts and the immense standing pool of microbial genetic variation will affect marine ecosystems' capacity to adjust to environmental changes. Here, we argue that significantly advancing our understanding of how host-associated microbes shape marine hosts' plastic and adaptive responses to environmental change requires (i) recognizing that individual host-microbe systems do not exist in an ecological or evolutionary vacuum and (ii) expanding the field toward long-term, multidisciplinary research on entire communities of hosts and microbes. Natural experiments, such as time-calibrated geological events associated with well-characterized environmental gradients, provide unique ecological and evolutionary contexts to address this challenge. We focus here particularly on mutualistic interactions between hosts and microbes, but note that many of the same lessons and approaches would apply to other types of interactions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
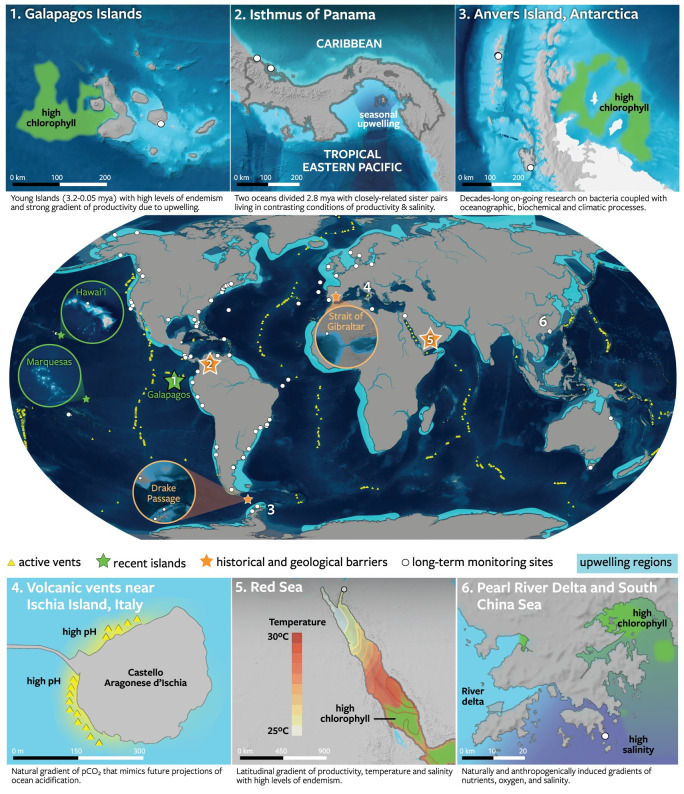
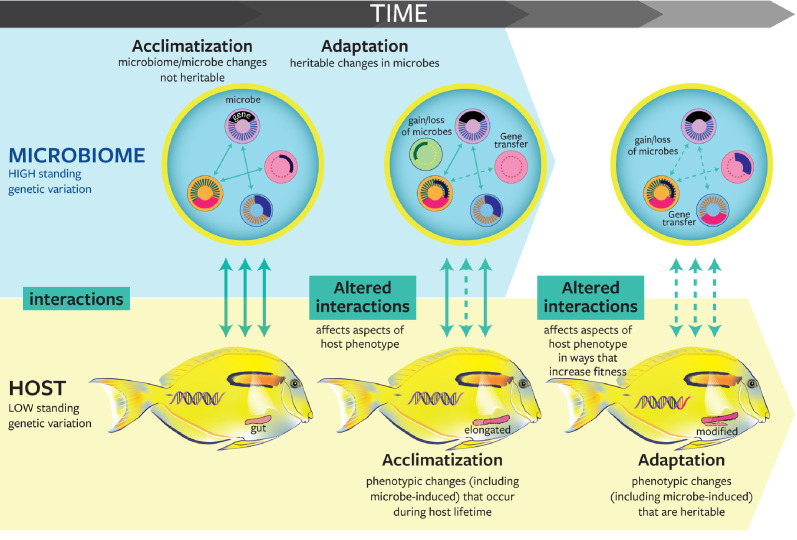
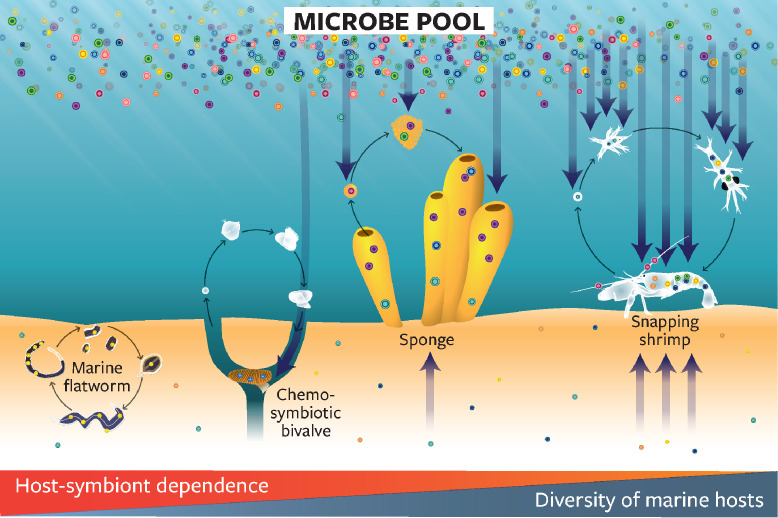
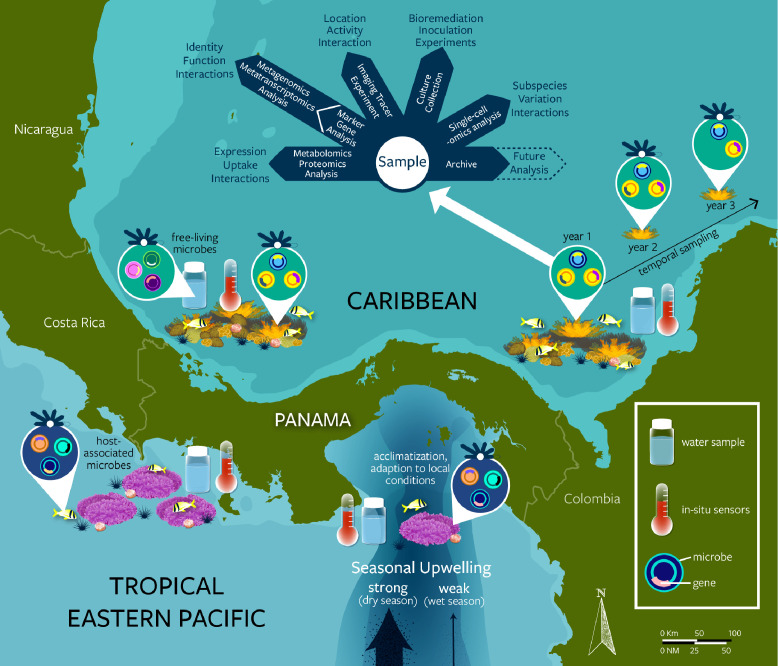
Similar articles
-
Host-associated microbiomes drive structure and function of marine ecosystems.PLoS Biol. 2019 Nov 11;17(11):e3000533. doi: 10.1371/journal.pbio.3000533. eCollection 2019 Nov. PLoS Biol. 2019. PMID: 31710600 Free PMC article.
-
Host control of the microbiome: Mechanisms, evolution, and disease.Science. 2024 Jul 19;385(6706):eadi3338. doi: 10.1126/science.adi3338. Epub 2024 Jul 19. Science. 2024. PMID: 39024451 Review.
-
Applying the core microbiome to understand host-microbe systems.J Anim Ecol. 2020 Jul;89(7):1549-1558. doi: 10.1111/1365-2656.13229. Epub 2020 Apr 14. J Anim Ecol. 2020. PMID: 32248522
-
Host-Microbe Coevolution: Applying Evidence from Model Systems to Complex Marine Invertebrate Holobionts.mBio. 2019 Feb 5;10(1):e02241-18. doi: 10.1128/mBio.02241-18. mBio. 2019. PMID: 30723123 Free PMC article. Review.
-
A place for host-microbe symbiosis in the comparative physiologist's toolbox.J Exp Biol. 2016 Nov 15;219(Pt 22):3496-3504. doi: 10.1242/jeb.136325. J Exp Biol. 2016. PMID: 27852759 Review.
Cited by
-
Preliminary study of shark microbiota at a unique mix-species shark aggregation site, in the Eastern Mediterranean Sea.Front Microbiol. 2023 Feb 23;14:1027804. doi: 10.3389/fmicb.2023.1027804. eCollection 2023. Front Microbiol. 2023. PMID: 36910211 Free PMC article.
-
Current knowledge of the Southern Hemisphere marine microbiome in eukaryotic hosts and the Strait of Magellan surface microbiome project.PeerJ. 2023 Oct 3;11:e15978. doi: 10.7717/peerj.15978. eCollection 2023. PeerJ. 2023. PMID: 37810788 Free PMC article. Review.
-
Microbial Community Dynamics Provide Evidence for Hypoxia during a Coral Reef Mortality Event.Appl Environ Microbiol. 2022 May 10;88(9):e0034722. doi: 10.1128/aem.00347-22. Epub 2022 Apr 18. Appl Environ Microbiol. 2022. PMID: 35435720 Free PMC article.
-
Contrasting drivers of abundant phage and prokaryotic communities revealed in diverse coastal ecosystems.ISME Commun. 2023 Dec 4;3(1):127. doi: 10.1038/s43705-023-00333-6. ISME Commun. 2023. PMID: 38049529 Free PMC article.
-
Biodiversity, Distribution and Functional Differences of Fungi in Four Species of Corals from the South China Sea, Elucidated by High-Throughput Sequencing Technology.J Fungi (Basel). 2024 Jun 27;10(7):452. doi: 10.3390/jof10070452. J Fungi (Basel). 2024. PMID: 39057337 Free PMC article.
