

Regulation of Citrobacter rodentium colonization: virulence, immune response and microbiota interactions
- PMID: 34352594
- PMCID: PMC8463437
- DOI: 10.1016/j.mib.2021.07.003
Regulation of Citrobacter rodentium colonization: virulence, immune response and microbiota interactions
Abstract
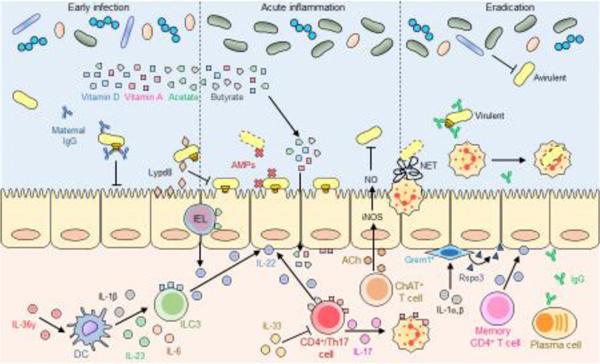
Citrobacter rodentium is a mouse-specific pathogen commonly used to model infection by human Enteropathogenic Escherichia coli, an important cause of infant diarrhea and mortality worldwide. In the early phase of infection, C. rodentium overcomes competition by the gut microbiota for successful replication. Then, the pathogen uses a type three secretion system (T3SS) to inject effector proteins into intestinal epithelial cells and induce metabolic and inflammatory conditions that promote colonization of the intestinal epithelium. C. rodentium also elicits highly coordinated innate and adaptive immune responses in the gut that regulate pathogen colonization and eradication. In this review, we highlight recent work on the regulation and function of the C. rodentium T3SS, the mechanisms employed by the pathogen to evade competition by the microbiota, and the function of the host immune response against infection.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures

Similar articles
-
Citrobacter rodentium: a model enteropathogen for understanding the interplay of innate and adaptive components of type 3 immunity.Mucosal Immunol. 2017 Sep;10(5):1108-1117. doi: 10.1038/mi.2017.47. Epub 2017 Jun 14. Mucosal Immunol. 2017. PMID: 28612839 Free PMC article. Review.
-
Citrobacter rodentium: infection, inflammation and the microbiota.Nat Rev Microbiol. 2014 Sep;12(9):612-23. doi: 10.1038/nrmicro3315. Epub 2014 Aug 4. Nat Rev Microbiol. 2014. PMID: 25088150 Review.
-
Linking Pathogen Virulence, Host Immunity and The Microbiota at the Intestinal Barrier.Keio J Med. 2017;66(1):14. doi: 10.2302/kjm.66-001-ABST. Keio J Med. 2017. PMID: 28356548
-
Modelling of infection by enteropathogenic Escherichia coli strains in lineages 2 and 4 ex vivo and in vivo by using Citrobacter rodentium expressing TccP.Infect Immun. 2009 Apr;77(4):1304-14. doi: 10.1128/IAI.01351-08. Epub 2009 Feb 2. Infect Immun. 2009. PMID: 19188355 Free PMC article.
-
The inhibition of COPII trafficking is important for intestinal epithelial tight junction disruption during enteropathogenic Escherichia coli and Citrobacter rodentium infection.Microbes Infect. 2013 Sep-Oct;15(10-11):738-44. doi: 10.1016/j.micinf.2013.05.001. Epub 2013 Jun 6. Microbes Infect. 2013. PMID: 23747681
Cited by
-
Simulated Colonic Fluid Replicates the In Vivo Growth Capabilities of Citrobacter rodentium cpxRA Mutants and Uncovers Additive Effects of Cpx-Regulated Genes on Fitness.Infect Immun. 2022 Sep 15;90(9):e0031422. doi: 10.1128/iai.00314-22. Epub 2022 Aug 24. Infect Immun. 2022. PMID: 36000875 Free PMC article.
-
Grimace scale assessment during Citrobacter rodentium inflammation and colitis development in laboratory mice.Front Vet Sci. 2023 Jun 5;10:1173446. doi: 10.3389/fvets.2023.1173446. eCollection 2023. Front Vet Sci. 2023. PMID: 37342621 Free PMC article.
-
Glucuronic acid confers colonization advantage to enteric pathogens.Proc Natl Acad Sci U S A. 2024 Mar 26;121(13):e2400226121. doi: 10.1073/pnas.2400226121. Epub 2024 Mar 19. Proc Natl Acad Sci U S A. 2024. PMID: 38502690 Free PMC article.
-
Edwardsiella piscicida infection reshapes the intestinal microbiome and metabolome of big-belly seahorses: mechanistic insights of synergistic actions of virulence factors.Front Immunol. 2023 May 3;14:1135588. doi: 10.3389/fimmu.2023.1135588. eCollection 2023. Front Immunol. 2023. PMID: 37215132 Free PMC article.
-
Akkermansia muciniphila Protects Against Antibiotic-Associated Diarrhea in Mice.Probiotics Antimicrob Proteins. 2024 Aug;16(4):1190-1204. doi: 10.1007/s12602-023-10101-6. Epub 2023 Jun 14. Probiotics Antimicrob Proteins. 2024. PMID: 37314693
References
-
- Collins JW, Keeney KM, Crepin VF, Rathinam VA, Fitzgerald KA, Finlay BB, Frankel G: Citrobacter rodentium: infection, inflammation and the microbiota. Nat Rev Microbiol 2014, 12:612–623. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
