

Transcriptional adaptation of Mycobacterium ulcerans in an original mouse model: New insights into the regulation of mycolactone
- PMID: 34107844
- PMCID: PMC8204960
- DOI: 10.1080/21505594.2021.1929749
Transcriptional adaptation of Mycobacterium ulcerans in an original mouse model: New insights into the regulation of mycolactone
Abstract
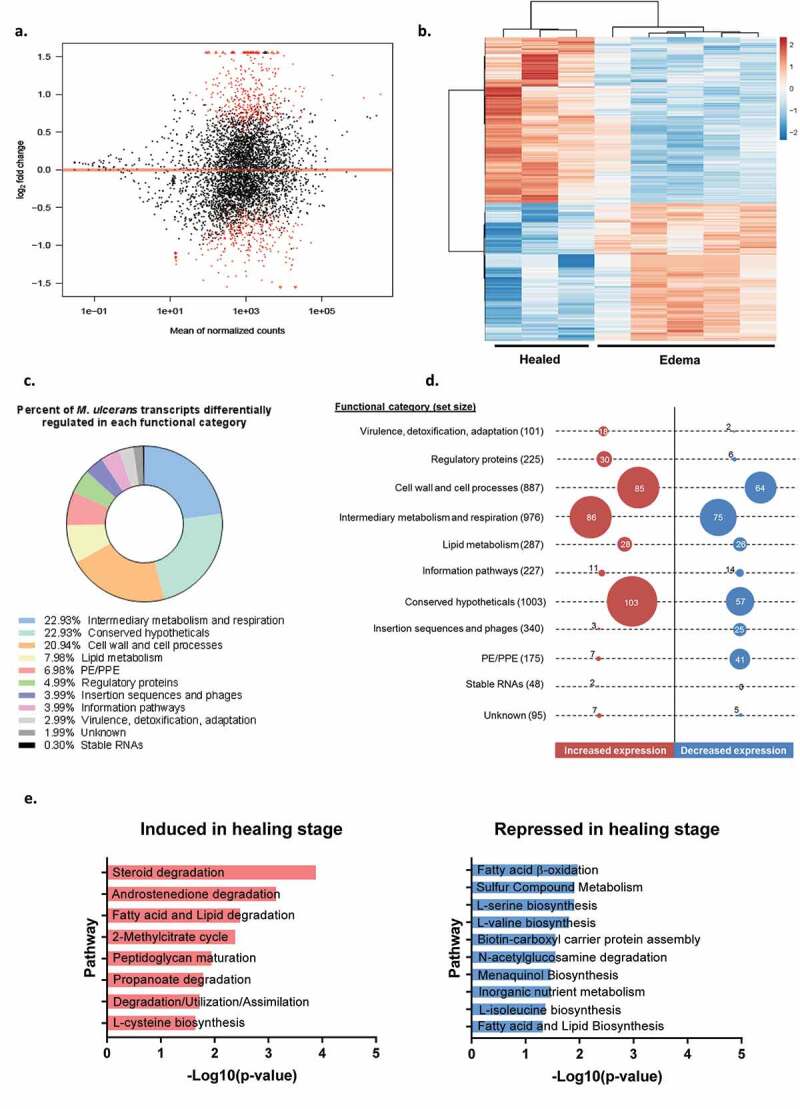
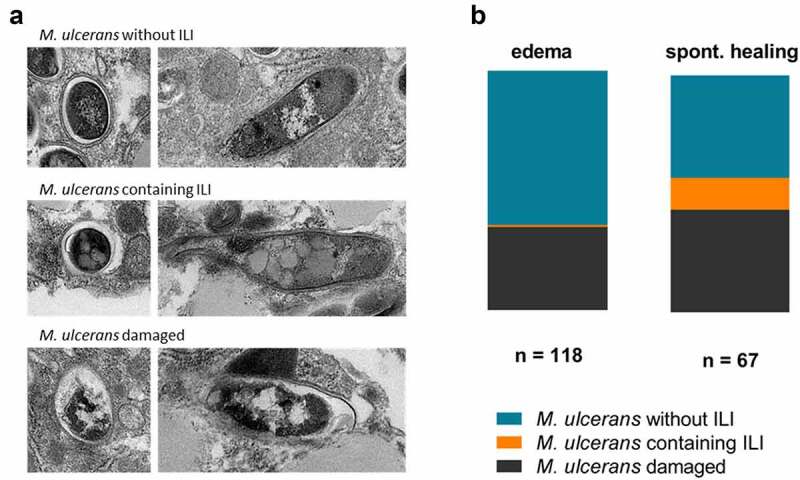
Mycobacterium ulcerans is the causal agent of Buruli ulcer, a chronic infectious disease and the third most common mycobacterial disease worldwide. Without early treatment, M. ulcerans provokes massive skin ulcers, caused by the mycolactone toxin, its main virulence factor. However, spontaneous healing may occur in Buruli ulcer patients several months or years after the disease onset. We have shown, in an original mouse model, that bacterial load remains high and viable in spontaneously healed tissues, with a switch of M. ulcerans to low levels of mycolactone production, adapting its strategy to survive in such a hostile environment. This original model offers the possibility to investigate the regulation of mycolactone production, by using an RNA-seq strategy to study bacterial adaptation during mouse infection. Pathway analysis and characterization of the tissue environment showed that the bacillus adapted to its new environment by modifying its metabolic activity and switching nutrient sources. Thus, M. ulcerans ensures its survival in healing tissues by reducing its secondary metabolism, leading to an inhibition of mycolactone synthesis. These findings shed new light on mycolactone regulation and pave the way for new therapeutic strategies.
Keywords: Mycobacterium ulcerans; host-bacterium interaction; metabolism; mycolactone; rna-sequencing.
Figures
Similar articles
-
FVB/N Mice Spontaneously Heal Ulcerative Lesions Induced by Mycobacterium ulcerans and Switch M. ulcerans into a Low Mycolactone Producer.J Immunol. 2016 Mar 15;196(6):2690-8. doi: 10.4049/jimmunol.1502194. Epub 2016 Feb 12. J Immunol. 2016. PMID: 26873988
-
Mycolactone gene expression is controlled by strong SigA-like promoters with utility in studies of Mycobacterium ulcerans and buruli ulcer.PLoS Negl Trop Dis. 2009 Nov 24;3(11):e553. doi: 10.1371/journal.pntd.0000553. PLoS Negl Trop Dis. 2009. PMID: 19936295 Free PMC article.
-
Skin-specific antibodies neutralizing mycolactone toxin during the spontaneous healing of Mycobacterium ulcerans infection.Sci Adv. 2020 Feb 26;6(9):eaax7781. doi: 10.1126/sciadv.aax7781. eCollection 2020 Feb. Sci Adv. 2020. PMID: 32133396 Free PMC article.
-
Immunity against Mycobacterium ulcerans: The subversive role of mycolactone.Immunol Rev. 2021 May;301(1):209-221. doi: 10.1111/imr.12956. Epub 2021 Feb 19. Immunol Rev. 2021. PMID: 33607704 Review.
-
Buruli ulcer and mycolactone-producing mycobacteria.Jpn J Infect Dis. 2013;66(2):83-8. doi: 10.7883/yoken.66.83. Jpn J Infect Dis. 2013. PMID: 23514902 Review.
Cited by
-
Repurposing of statins for Buruli Ulcer treatment: antimicrobial activity against Mycobacterium ulcerans.Front Microbiol. 2023 Sep 29;14:1266261. doi: 10.3389/fmicb.2023.1266261. eCollection 2023. Front Microbiol. 2023. PMID: 37840746 Free PMC article.
-
Induced Synthesis of Mycolactone Restores the Pathogenesis of Mycobacterium ulcerans In Vitro and In Vivo.Front Immunol. 2022 Mar 24;13:750643. doi: 10.3389/fimmu.2022.750643. eCollection 2022. Front Immunol. 2022. PMID: 35401531 Free PMC article.
-
Genome-wide screening identified SEC61A1 as an essential factor for mycolactone-dependent apoptosis in human premonocytic THP-1 cells.PLoS Negl Trop Dis. 2022 Aug 8;16(8):e0010672. doi: 10.1371/journal.pntd.0010672. eCollection 2022 Aug. PLoS Negl Trop Dis. 2022. PMID: 35939511 Free PMC article.
-
Intrabacterial lipid inclusion-associated proteins: a core machinery conserved from saprophyte Actinobacteria to the human pathogen Mycobacterium tuberculosis.FEBS Open Bio. 2023 Dec;13(12):2306-2323. doi: 10.1002/2211-5463.13721. Epub 2023 Nov 15. FEBS Open Bio. 2023. PMID: 37872001 Free PMC article.
-
The One That Got Away: How Macrophage-Derived IL-1β Escapes the Mycolactone-Dependent Sec61 Blockade in Buruli Ulcer.Front Immunol. 2022 Jan 26;12:788146. doi: 10.3389/fimmu.2021.788146. eCollection 2021. Front Immunol. 2022. PMID: 35154073 Free PMC article.
References
-
- Portaels F, Silva MT, Meyers WM.. Buruli ulcer. Clin Dermatol. 2009;27(3):291–305. - PubMed
-
- Barogui Y, Johnson RC, Van Der Werf TS, et al. Functional limitations after surgical or antibiotic treatment for Buruli ulcer in Benin. Am J Trop Med Hyg. 2009;81(1):82–87. - PubMed
-
- Vincent QB, Ardant MF, Adeye A, et al. Clinical epidemiology of laboratory-confirmed Buruli ulcer in Benin: a cohort study. Lancet Glob Health. 2014;2(7):e422–30. - PubMed
-
- Wadagni AC, Barogui YT, Johnson RC, et al. Delayed versus standard assessment for excision surgery in patients with Buruli ulcer in Benin: a randomised controlled trial. Lancet Infect Dis. 2018;18(6):650–656. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases