

Splicing and editing of ionotropic glutamate receptors: a comprehensive analysis based on human RNA-Seq data
- PMID: 34100982
- PMCID: PMC8257547
- DOI: 10.1007/s00018-021-03865-z
Splicing and editing of ionotropic glutamate receptors: a comprehensive analysis based on human RNA-Seq data
Abstract
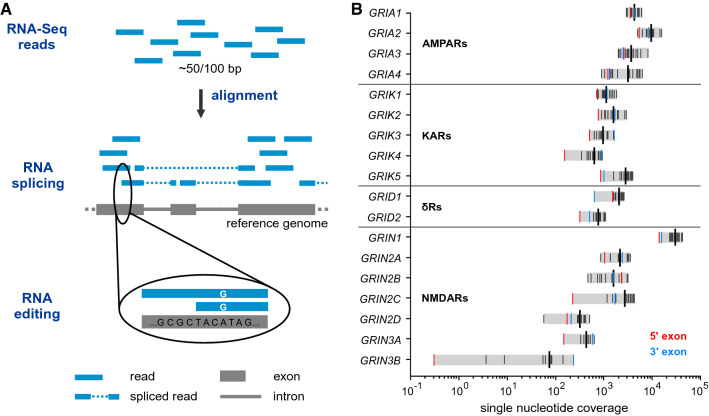
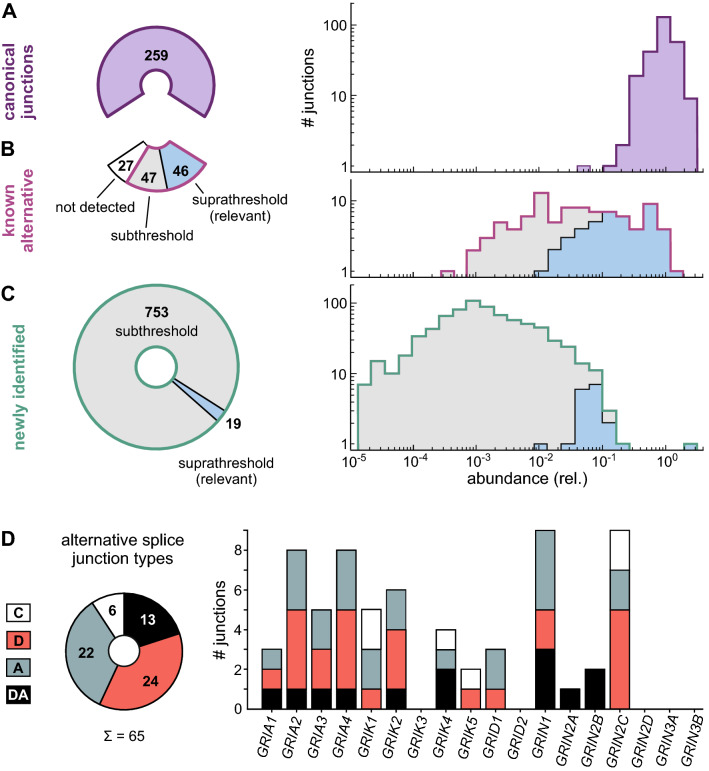
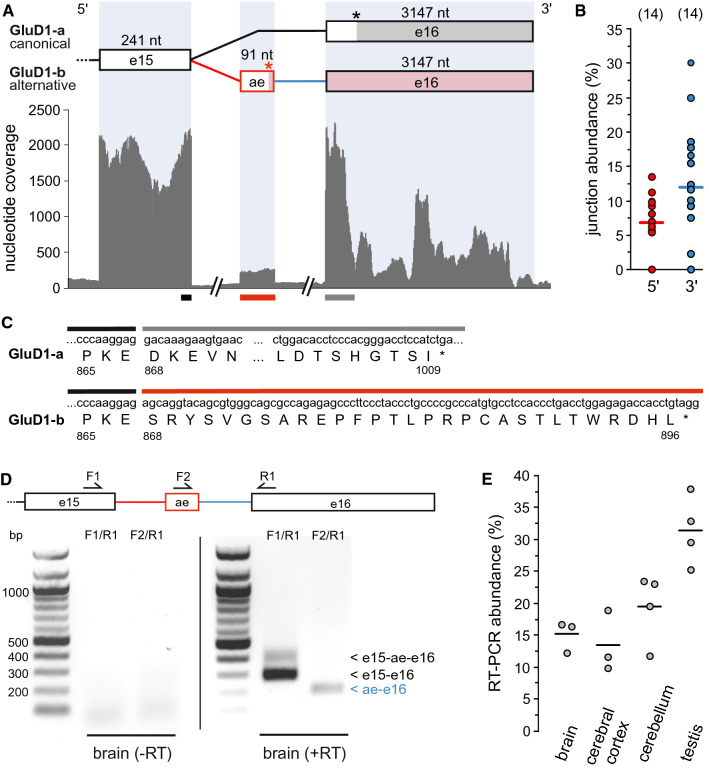
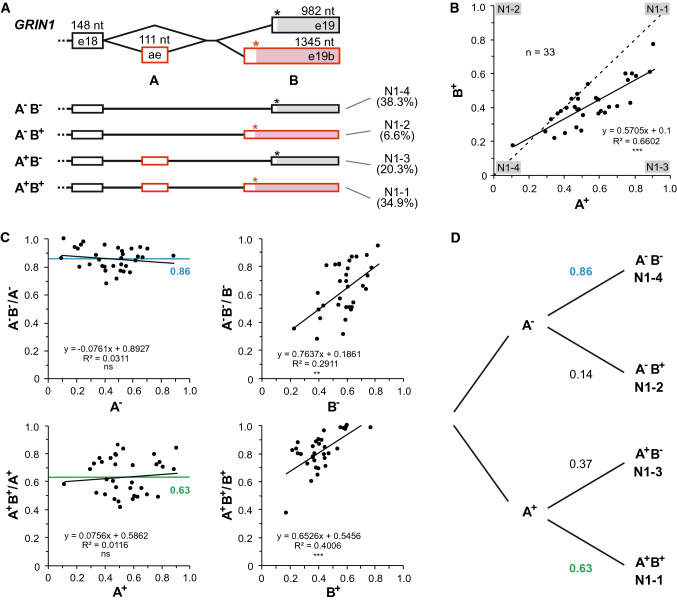
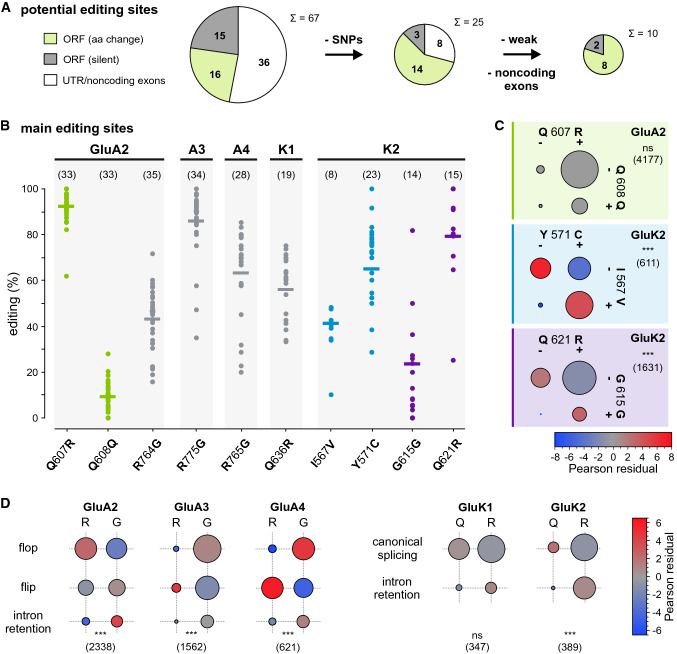
Ionotropic glutamate receptors (iGluRs) play key roles for signaling in the central nervous system. Alternative splicing and RNA editing are well-known mechanisms to increase iGluR diversity and to provide context-dependent regulation. Earlier work on isoform identification has focused on the analysis of cloned transcripts, mostly from rodents. We here set out to obtain a systematic overview of iGluR splicing and editing in human brain based on RNA-Seq data. Using data from two large-scale transcriptome studies, we established a workflow for the de novo identification and quantification of alternative splice and editing events. We detected all canonical iGluR splice junctions, assessed the abundance of alternative events described in the literature, and identified new splice events in AMPA, kainate, delta, and NMDA receptor subunits. Notable events include an abundant transcript encoding the GluA4 amino-terminal domain, GluA4-ATD, a novel C-terminal GluD1 (delta receptor 1) isoform, GluD1-b, and potentially new GluK4 and GluN2C isoforms. C-terminal GluN1 splicing may be controlled by inclusion of a cassette exon, which shows preference for one of the two acceptor sites in the last exon. Moreover, we identified alternative untranslated regions (UTRs) and species-specific differences in splicing. In contrast, editing in exonic iGluR regions appears to be mostly limited to ten previously described sites, two of which result in silent amino acid changes. Coupling of proximal editing/editing and editing/splice events occurs to variable degree. Overall, this analysis provides the first inventory of alternative splicing and editing in human brain iGluRs and provides the impetus for further transcriptome-based and functional investigations.
Keywords: C-to-U editing; Next-generation sequencing (NGS); Nonsense-mediated decay (NMD); Primate-specific; Single-nucleotide polymorphism (SNP); Splicing error.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
Similar articles
-
Ionotropic glutamate receptor mRNA editing in the prefrontal cortex: no alterations in schizophrenia or bipolar disorder.J Psychiatry Neurosci. 2012 Jul;37(4):267-72. doi: 10.1503/jpn.110107. J Psychiatry Neurosci. 2012. PMID: 22469055 Free PMC article.
-
AMPA, NMDA and kainate glutamate receptor subunits are expressed in human peripheral blood mononuclear cells (PBMCs) where the expression of GluK4 is altered by pregnancy and GluN2D by depression in pregnant women.J Neuroimmunol. 2017 Apr 15;305:51-58. doi: 10.1016/j.jneuroim.2017.01.013. Epub 2017 Jan 21. J Neuroimmunol. 2017. PMID: 28284346
-
Glutamate receptor RNA editing in health and disease.Biochemistry (Mosc). 2011 Aug;76(8):882-9. doi: 10.1134/S0006297911080037. Biochemistry (Mosc). 2011. PMID: 22022961 Review.
-
Automated Isoform Diversity Detector (AIDD): a pipeline for investigating transcriptome diversity of RNA-seq data.BMC Bioinformatics. 2020 Dec 30;21(Suppl 18):578. doi: 10.1186/s12859-020-03888-6. BMC Bioinformatics. 2020. PMID: 33375933 Free PMC article.
-
Multiplexed primer extension sequencing: A targeted RNA-seq method that enables high-precision quantitation of mRNA splicing isoforms and rare pre-mRNA splicing intermediates.Methods. 2020 Apr 1;176:34-45. doi: 10.1016/j.ymeth.2019.05.013. Epub 2019 May 21. Methods. 2020. PMID: 31121301 Free PMC article. Review.
Cited by
-
Temporal landscape and translational regulation of A-to-I RNA editing in mouse retina development.BMC Biol. 2024 May 7;22(1):106. doi: 10.1186/s12915-024-01908-y. BMC Biol. 2024. PMID: 38715001 Free PMC article.
-
Hippocampal adenosine-to-inosine RNA editing in sepsis: dynamic changes and influencing factors.Brain Commun. 2024 Aug 8;6(4):fcae260. doi: 10.1093/braincomms/fcae260. eCollection 2024. Brain Commun. 2024. PMID: 39135964 Free PMC article.
-
Calcium- and calmodulin-dependent inhibition of NMDA receptor currents.Biophys J. 2024 Feb 6;123(3):277-293. doi: 10.1016/j.bpj.2023.12.018. Epub 2023 Dec 22. Biophys J. 2024. PMID: 38140727 Review.
-
Preferential potentiation of AMPA-mediated currents in brainstem hypoglossal motoneurons by subchronic exposure of mice expressing the human superoxide dismutase 1 G93A gene mutation to neurotoxicant methylmercury in vivo.Neurotoxicology. 2024 Jan;100:72-84. doi: 10.1016/j.neuro.2023.12.002. Epub 2023 Dec 7. Neurotoxicology. 2024. PMID: 38065418 Free PMC article.
-
Genome-wide epigenetic modifications in sports horses during training as an adaptation phenomenon.Sci Rep. 2023 Nov 1;13(1):18786. doi: 10.1038/s41598-023-46043-w. Sci Rep. 2023. PMID: 37914824 Free PMC article.
References
-
- Saito Y, Yuan Y, Zucker-Scharff I, Fak JJ, Jereb S, Tajima Y, Licatalosi DD, Darnell RB. Differential NOVA2-mediated splicing in excitatory and inhibitory neurons regulates cortical development and cerebellar function. Neuron. 2019;101:707–720.e705. doi: 10.1016/j.neuron.2018.12.019. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
