

Effective Drug Concentration and Selectivity Depends on Fraction of Primitive Cells
- PMID: 34066491
- PMCID: PMC8125035
- DOI: 10.3390/ijms22094931
Effective Drug Concentration and Selectivity Depends on Fraction of Primitive Cells
Abstract
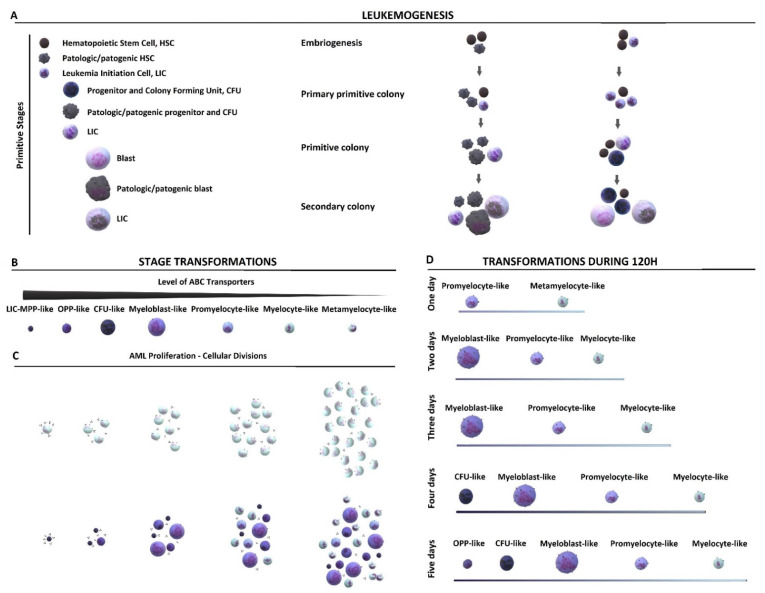
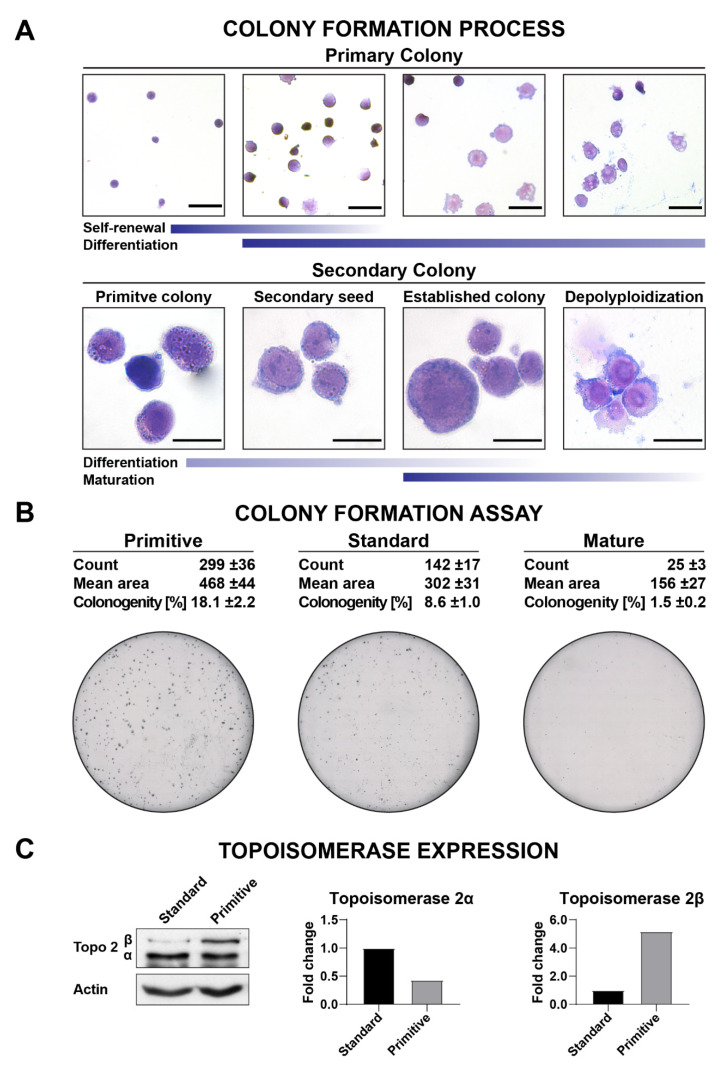
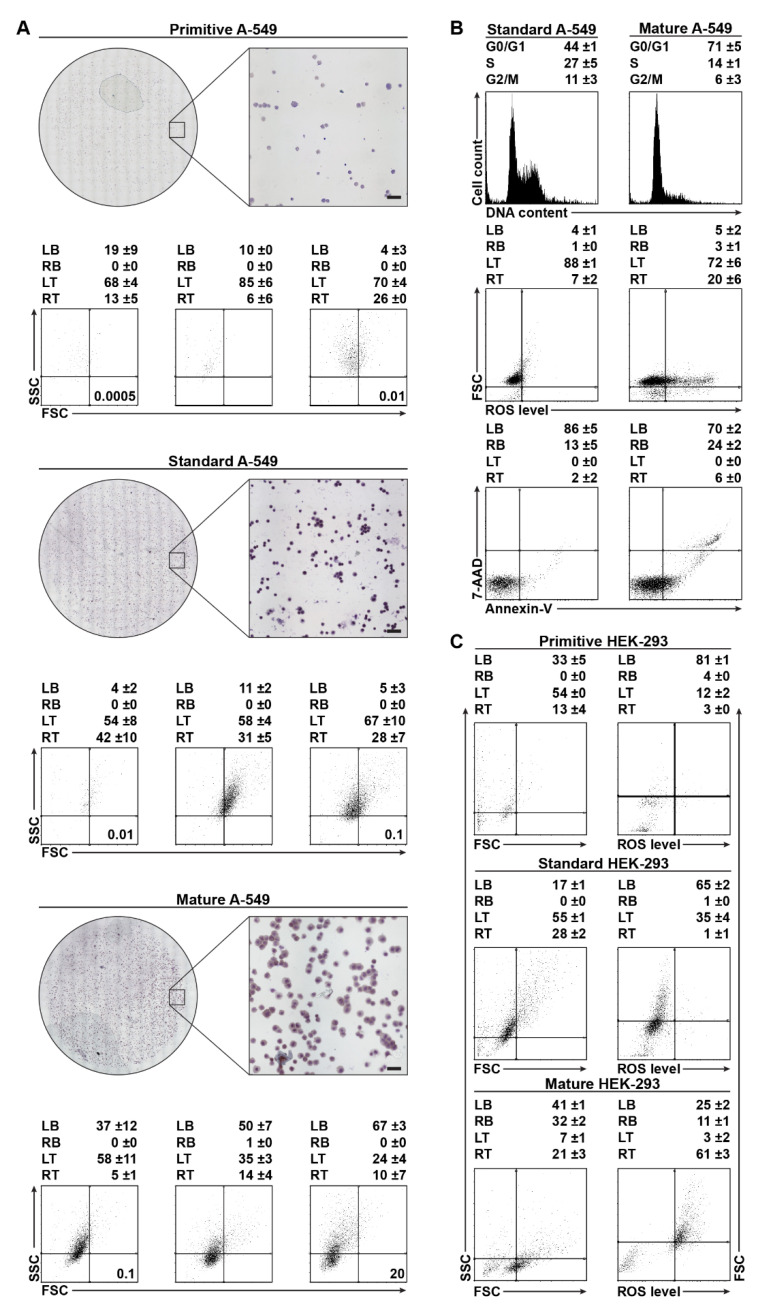
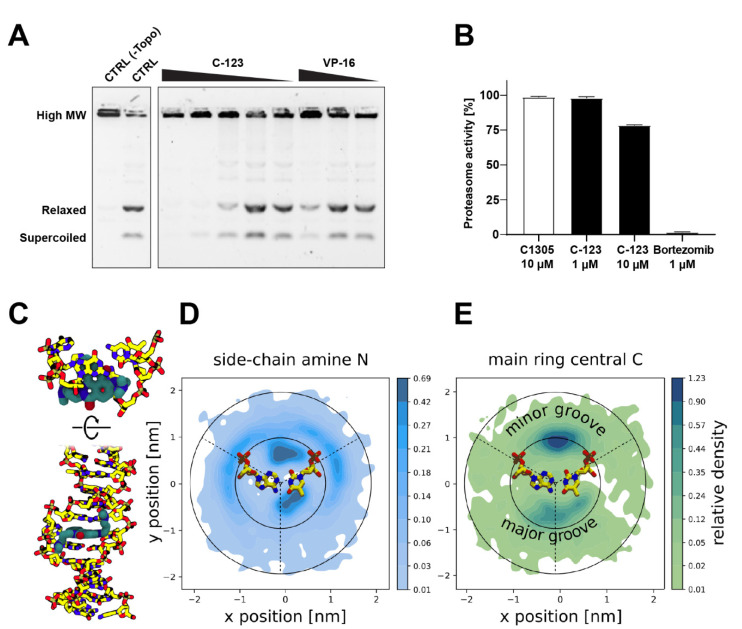
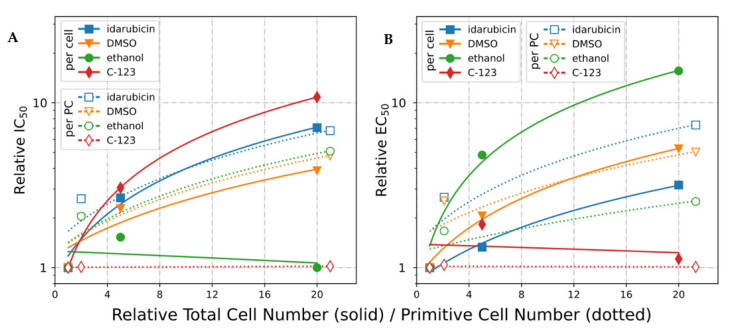
Poor efficiency of chemotherapeutics in the eradication of Cancer Stem Cells (CSCs) has been driving the search for more active and specific compounds. In this work, we show how cell density-dependent stage culture profiles can be used in drug development workflows to achieve more robust drug activity (IC50 and EC50) results. Using flow cytometry and light microscopy, we characterized the cytological stage profiles of the HL-60-, A-549-, and HEK-293-derived sublines with a focus on their primitive cell content. We then used a range of cytotoxic substances-C-123, bortezomib, idarubicin, C-1305, doxorubicin, DMSO, and ethanol-to highlight typical density-related issues accompanying drug activity determination. We also showed that drug EC50 and selectivity indices normalized to primitive cell content are more accurate activity measurements. We tested our approach by calculating the corrected selectivity index of a novel chemotherapeutic candidate, C-123. Overall, our study highlights the usefulness of accounting for primitive cell fractions in the assessment of drug efficiency.
Keywords: C-123; cell culture density-dependent cytological stage profile; drug screening platform; effective drug concentration; primitive acute myeloid leukemia cellular stages; primitive cancer cellular stages; selectivity index.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Cell Density-Dependent Cytological Stage Profile and Its Application for a Screen of Cytostatic Agents Active Toward Leukemic Stem Cells.Stem Cells Dev. 2018 Apr 1;27(7):488-513. doi: 10.1089/scd.2017.0245. Epub 2018 Mar 13. Stem Cells Dev. 2018. PMID: 29431006
-
Reactive oxygen species mediate cancer stem-like cells and determine bortezomib sensitivity via Mcl-1 and Zeb-1 in mantle cell lymphoma.Biochim Biophys Acta Mol Basis Dis. 2018 Nov;1864(11):3739-3753. doi: 10.1016/j.bbadis.2018.09.010. Epub 2018 Sep 11. Biochim Biophys Acta Mol Basis Dis. 2018. PMID: 30251692
-
Salinomycin induces apoptosis in cisplatin-resistant colorectal cancer cells by accumulation of reactive oxygen species.Toxicol Lett. 2013 Oct 24;222(2):139-45. doi: 10.1016/j.toxlet.2013.07.022. Epub 2013 Aug 2. Toxicol Lett. 2013. PMID: 23916687
-
Primitive Cancer Cell States: A Target for Drug Screening?Trends Pharmacol Sci. 2019 Mar;40(3):161-171. doi: 10.1016/j.tips.2019.01.003. Epub 2019 Jan 29. Trends Pharmacol Sci. 2019. PMID: 30709543 Review.
-
Quantitative cytology in leukemia research.Blood Cells. 1983;9(1):35-55. Blood Cells. 1983. PMID: 6344934 Review.
Cited by
-
Inducing Cytotoxicity in Colon Cancer Cells and Suppressing Cancer Stem Cells by Dolasetron and Ketoprofen through Inhibition of RNA Binding Protein PUM1.Toxics. 2023 Aug 3;11(8):669. doi: 10.3390/toxics11080669. Toxics. 2023. PMID: 37624174 Free PMC article.
-
Antiviral Activity of trans-Hexenoic Acid against Coxsackievirus B and Enterovirus A71.Antimicrob Agents Chemother. 2023 Mar 16;67(3):e0086822. doi: 10.1128/aac.00868-22. Epub 2023 Feb 14. Antimicrob Agents Chemother. 2023. PMID: 36786598 Free PMC article.
-
Three New Dihydrophenanthrene Derivatives from Cymbidium ensifolium and Their Cytotoxicity against Cancer Cells.Molecules. 2022 Mar 29;27(7):2222. doi: 10.3390/molecules27072222. Molecules. 2022. PMID: 35408617 Free PMC article.
-
Molecularly Engineered Surfactin Analogues Induce Nonapoptotic-Like Cell Death and Increased Selectivity in Multiple Breast Cancer Cell Types.ACS Omega. 2023 Apr 10;8(16):14610-14620. doi: 10.1021/acsomega.3c00454. eCollection 2023 Apr 25. ACS Omega. 2023. PMID: 37125141 Free PMC article.
-
(Very) Small Stem-like Cells in Human Cell Cultures.Cancers (Basel). 2023 Nov 22;15(23):5520. doi: 10.3390/cancers15235520. Cancers (Basel). 2023. PMID: 38067224 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
