

Cellular functions of actin- and microtubule-associated septins
- PMID: 34033796
- PMCID: PMC8194058
- DOI: 10.1016/j.cub.2021.03.064
Cellular functions of actin- and microtubule-associated septins
Abstract
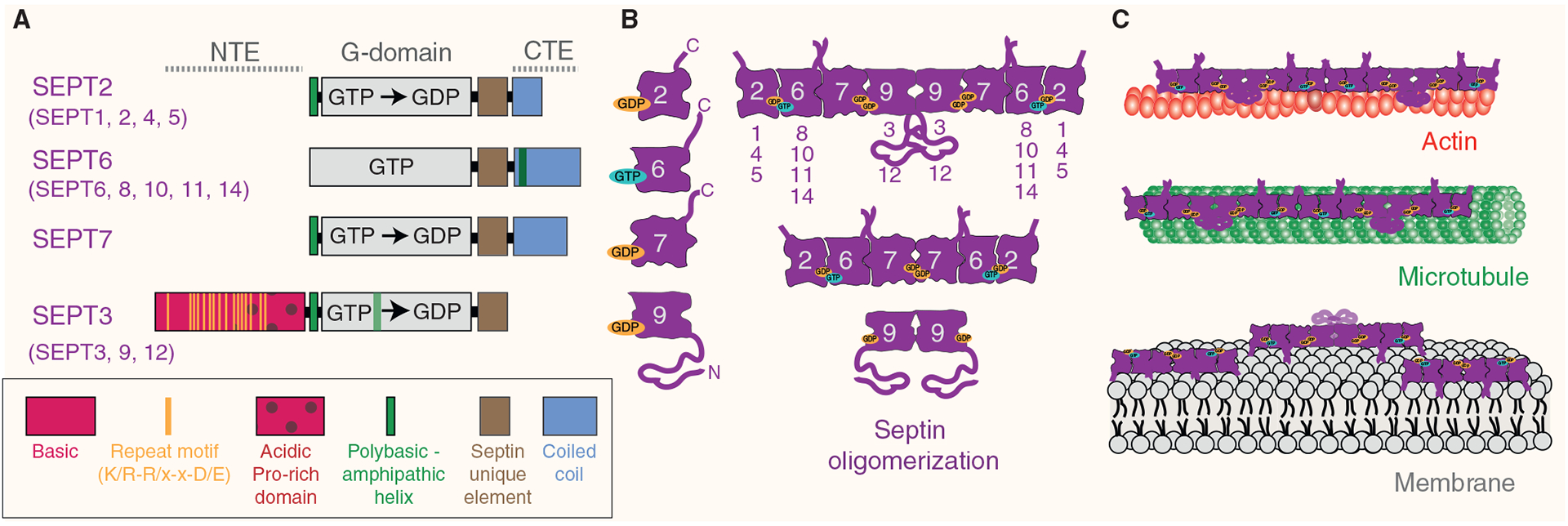
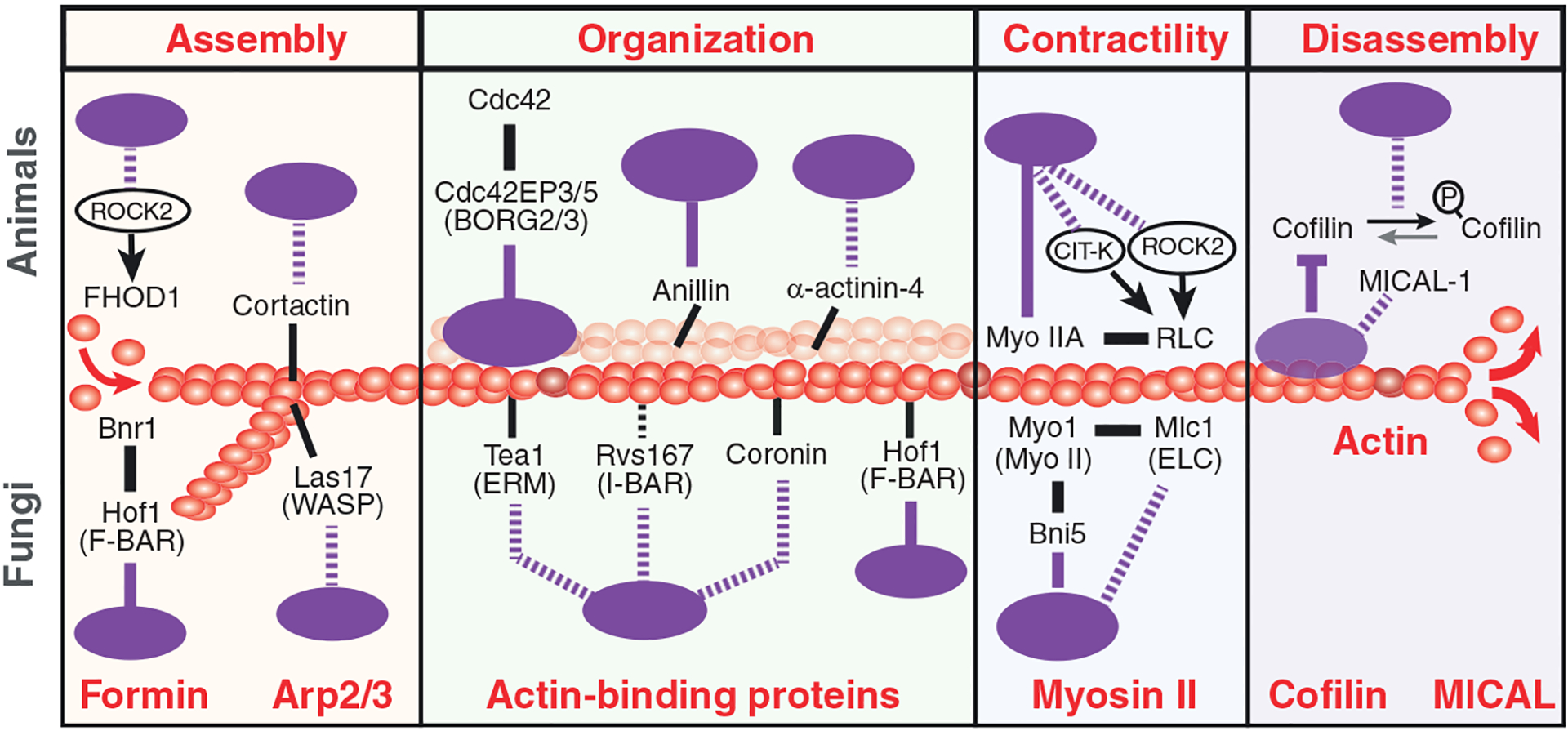
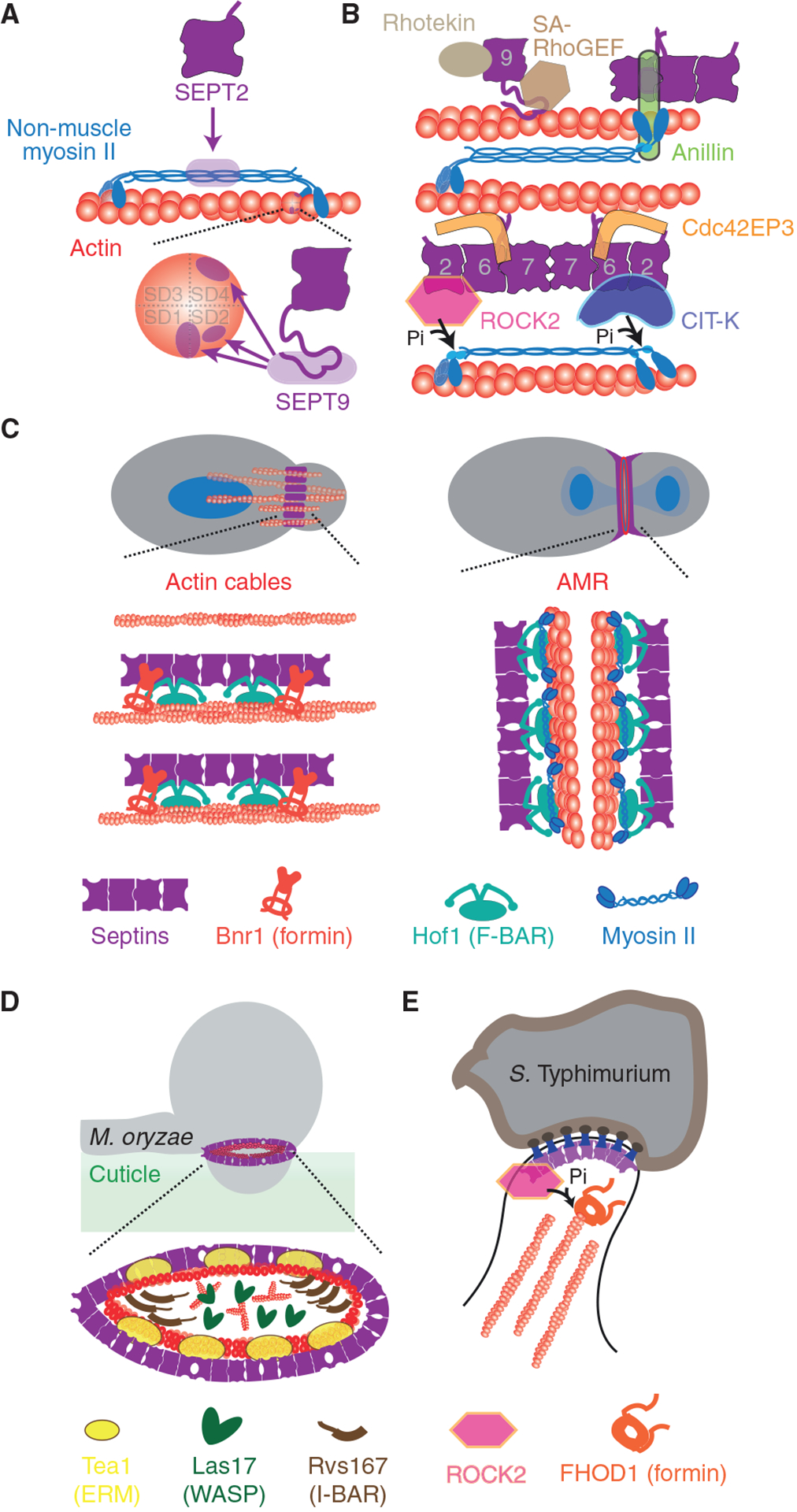
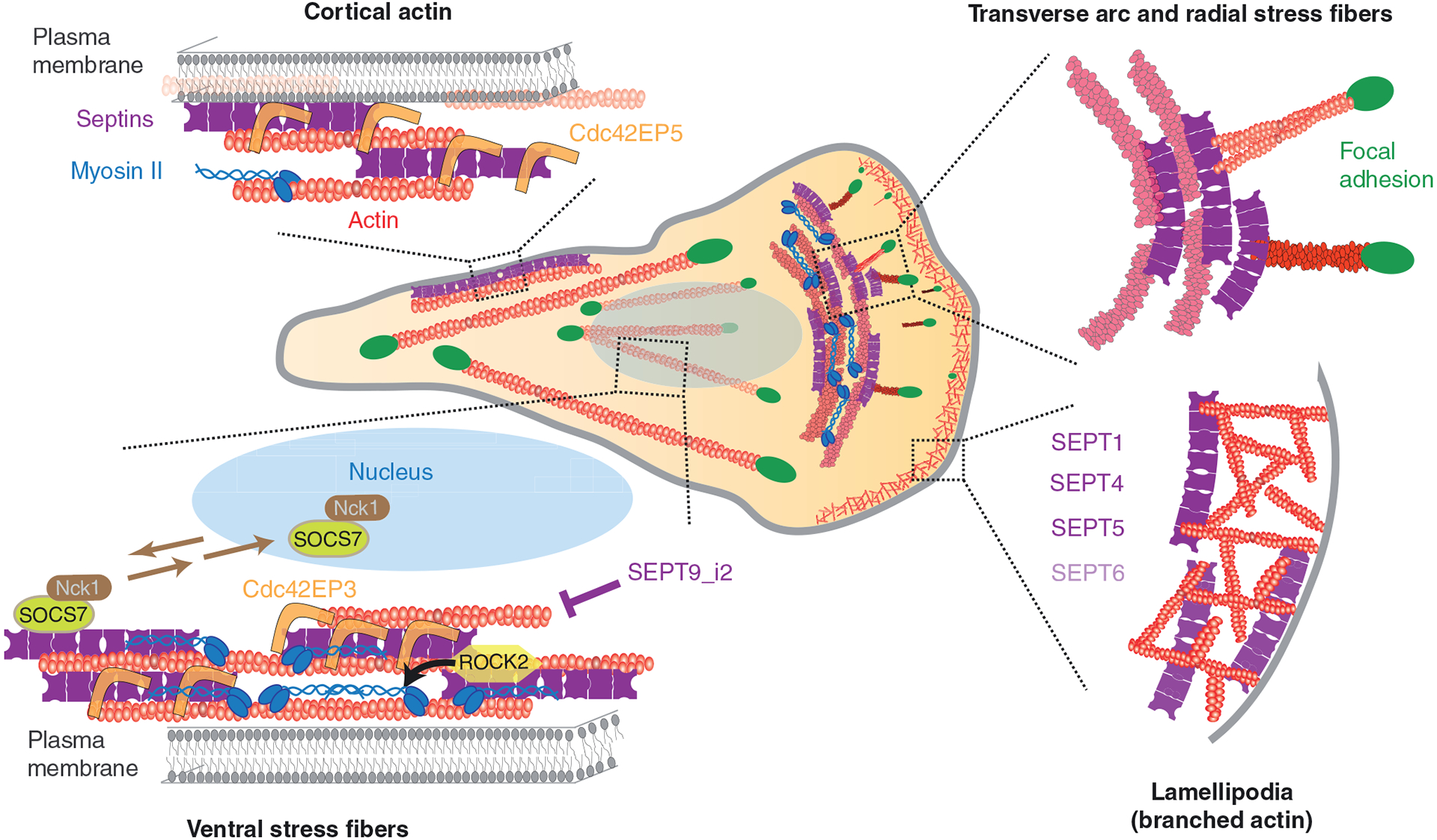
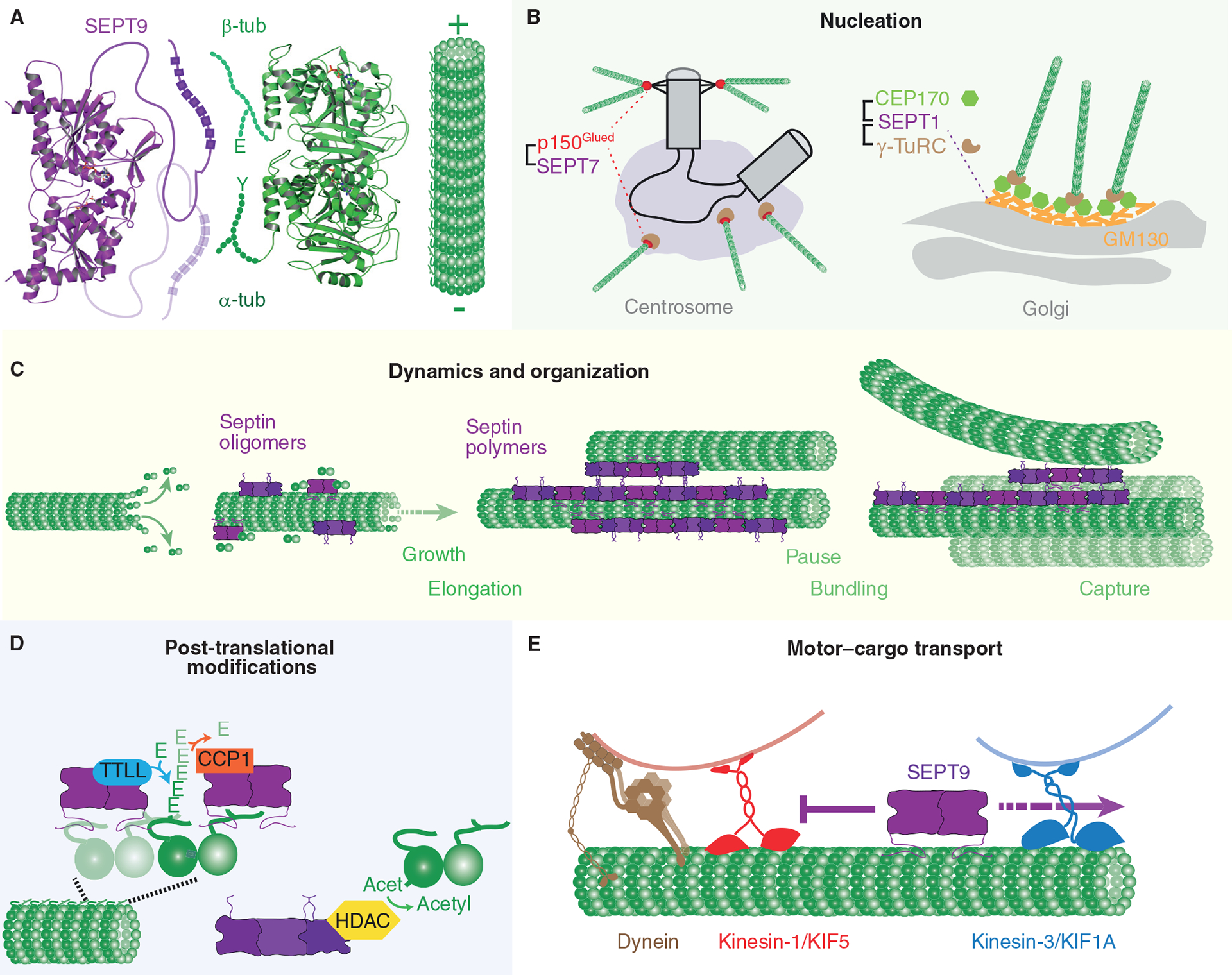
Septins are an integral component of the cytoskeleton, assembling into higher-order oligomers and filamentous polymers that associate with actin filaments, microtubules and membranes. Here, we review septin interactions with actin and microtubules, and septin-mediated regulation of the organization and dynamics of these cytoskeletal networks, which is critical for cellular morphogenesis. We discuss how actomyosin-associated septins function in cytokinesis, cell migration and host defense against pathogens. We highlight newly emerged roles of septins at the interface of microtubules and membranes with molecular motors, which point to a 'septin code' for the regulation of membrane traffic. Additionally, we revisit the functions of microtubule-associated septins in mitosis and meiosis. In sum, septins comprise a unique module of cytoskeletal regulators that are spatially and functionally specialized and have properties of bona fide actin-binding and microtubule-associated proteins. With many questions still outstanding, the study of septins will continue to provide new insights into fundamental problems of cytoskeletal organization and function.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Spatial effects - site-specific regulation of actin and microtubule organization by septin GTPases.J Cell Sci. 2018 Jan 11;131(1):jcs207555. doi: 10.1242/jcs.207555. J Cell Sci. 2018. PMID: 29326311 Free PMC article. Review.
-
Septins mediate a microtubule-actin crosstalk that enables actin growth on microtubules.Proc Natl Acad Sci U S A. 2022 Dec 13;119(50):e2202803119. doi: 10.1073/pnas.2202803119. Epub 2022 Dec 7. Proc Natl Acad Sci U S A. 2022. PMID: 36475946 Free PMC article.
-
Masters of asymmetry - lessons and perspectives from 50 years of septins.Mol Biol Cell. 2020 Oct 1;31(21):2289-2297. doi: 10.1091/mbc.E19-11-0648. Mol Biol Cell. 2020. PMID: 32991244 Free PMC article. Review.
-
Septins promote F-actin ring formation by crosslinking actin filaments into curved bundles.Nat Cell Biol. 2014 Apr;16(4):322-34. doi: 10.1038/ncb2921. Epub 2014 Mar 16. Nat Cell Biol. 2014. PMID: 24633326
-
Actin, microtubule, septin and ESCRT filament remodeling during late steps of cytokinesis.Curr Opin Cell Biol. 2018 Feb;50:27-34. doi: 10.1016/j.ceb.2018.01.007. Epub 2018 Feb 10. Curr Opin Cell Biol. 2018. PMID: 29438904 Review.
Cited by
-
Establishing neuronal polarity: microtubule regulation during neurite initiation.Oxf Open Neurosci. 2022 May 13;1:kvac007. doi: 10.1093/oons/kvac007. eCollection 2022. Oxf Open Neurosci. 2022. PMID: 38596701 Free PMC article. Review.
-
Septin-coated microtubules promote maturation of multivesicular bodies by inhibiting their motility.J Cell Biol. 2024 Aug 5;223(8):e202308049. doi: 10.1083/jcb.202308049. Epub 2024 Apr 26. J Cell Biol. 2024. PMID: 38668767 Free PMC article.
-
Dynamic remodeling of septin structures fine-tunes myogenic differentiation.iScience. 2024 Jul 31;27(9):110630. doi: 10.1016/j.isci.2024.110630. eCollection 2024 Sep 20. iScience. 2024. PMID: 39246450 Free PMC article.
-
Pyramidal neuron morphogenesis requires a septin network that stabilizes filopodia and suppresses lamellipodia during neurite initiation.Curr Biol. 2023 Feb 6;33(3):434-448.e8. doi: 10.1016/j.cub.2022.11.043. Epub 2022 Dec 19. Curr Biol. 2023. PMID: 36538929 Free PMC article.
-
The Structural Biology of Septins and Their Filaments: An Update.Front Cell Dev Biol. 2021 Nov 19;9:765085. doi: 10.3389/fcell.2021.765085. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34869357 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
