

How epigallocatechin gallate binds and assembles oligomeric forms of human alpha-synuclein
- PMID: 34019875
- PMCID: PMC8191297
- DOI: 10.1016/j.jbc.2021.100788
How epigallocatechin gallate binds and assembles oligomeric forms of human alpha-synuclein
Abstract
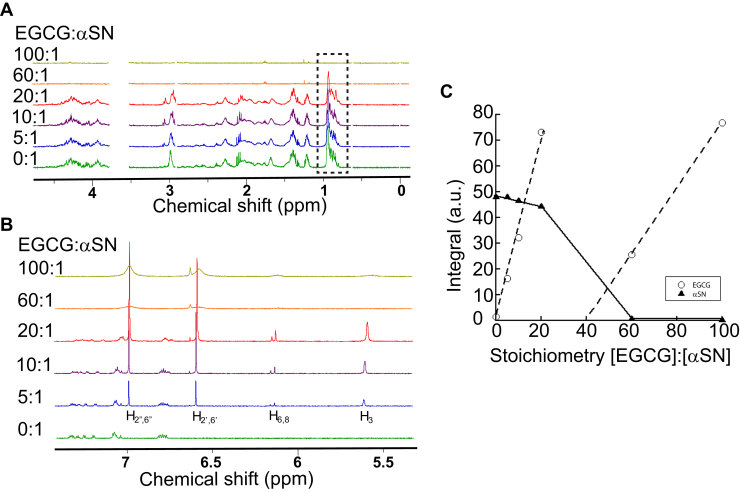
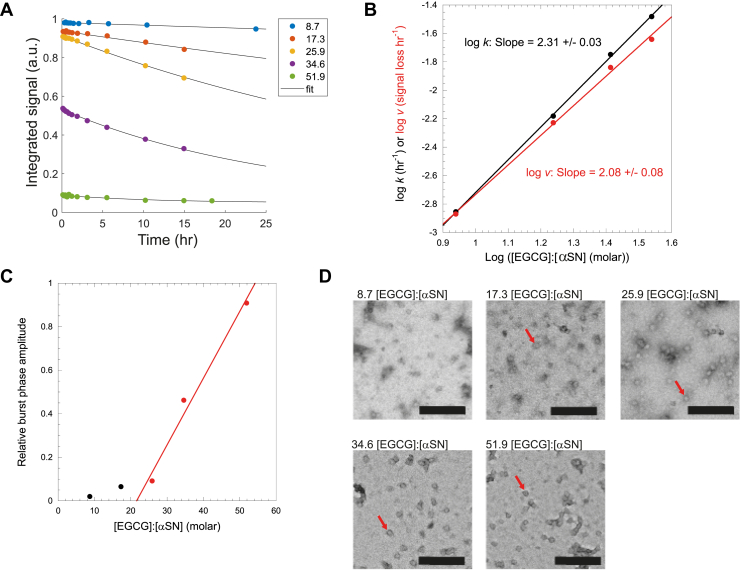
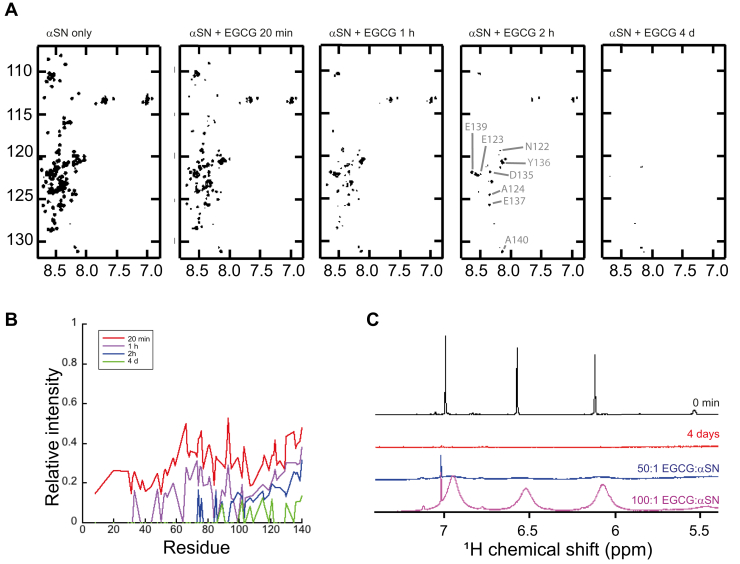
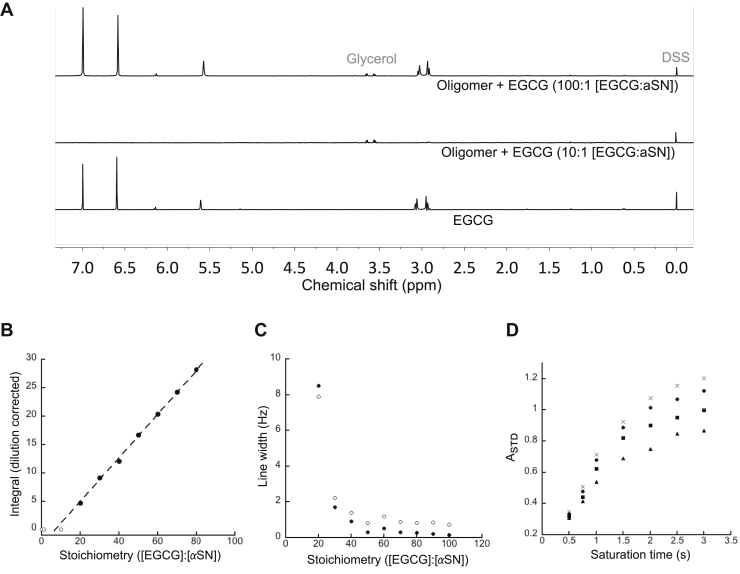
The intrinsically disordered human protein α-synuclein (αSN) can self-associate into oligomers and amyloid fibrils. Several lines of evidence suggest that oligomeric αSN is cytotoxic, making it important to devise strategies to either prevent oligomer formation and/or inhibit the ensuing toxicity. (-)-epigallocatechin gallate (EGCG) has emerged as a molecular modulator of αSN self-assembly, as it reduces the flexibility of the C-terminal region of αSN in the oligomer and inhibits the oligomer's ability to perturb phospholipid membranes and induce cell death. However, a detailed structural and kinetic characterization of this interaction is still lacking. Here, we use liquid-state NMR spectroscopy to investigate how EGCG interacts with monomeric and oligomeric forms of αSN. We find that EGCG can bind to all parts of monomeric αSN but exhibits highest affinity for the N-terminal region. Monomeric αSN binds ∼54 molecules of EGCG in total during oligomerization. Furthermore, kinetic data suggest that EGCG dimerization is coupled with the αSN association reaction. In contrast, preformed oligomers only bind ∼7 EGCG molecules per protomer, in agreement with the more compact nature of the oligomer compared with the natively unfolded monomer. In previously conducted cell assays, as little as 0.36 EGCG per αSN reduce oligomer toxicity by 50%. Our study thus demonstrates that αSN cytotoxicity can be inhibited by small molecules at concentrations at least an order of magnitude below full binding capacity. We speculate this is due to cooperative binding of protein-stabilized EGCG dimers, which in turn implies synergy between protein association and EGCG dimerization.
Keywords: NMR; alpha-synuclein (α-synuclein); kinetics; oligomer; oligomerization; protein aggregation; stoichiometry.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Similar articles
-
How epigallocatechin gallate can inhibit α-synuclein oligomer toxicity in vitro.J Biol Chem. 2014 Aug 1;289(31):21299-310. doi: 10.1074/jbc.M114.554667. Epub 2014 Jun 6. J Biol Chem. 2014. PMID: 24907278 Free PMC article.
-
Potent α-Synuclein Aggregation Inhibitors, Identified by High-Throughput Screening, Mainly Target the Monomeric State.Cell Chem Biol. 2018 Nov 15;25(11):1389-1402.e9. doi: 10.1016/j.chembiol.2018.08.005. Epub 2018 Sep 6. Cell Chem Biol. 2018. PMID: 30197194
-
The N-terminus of α-synuclein is essential for both monomeric and oligomeric interactions with membranes.FEBS Lett. 2014 Jan 31;588(3):497-502. doi: 10.1016/j.febslet.2013.12.015. Epub 2013 Dec 25. FEBS Lett. 2014. PMID: 24374342
-
Phase separation and other forms of α-Synuclein self-assemblies.Essays Biochem. 2022 Dec 16;66(7):987-1000. doi: 10.1042/EBC20220055. Essays Biochem. 2022. PMID: 36373662 Review.
-
Green Tea Epigallocatechin-3-gallate (EGCG) Targeting Protein Misfolding in Drug Discovery for Neurodegenerative Diseases.Biomolecules. 2021 May 20;11(5):767. doi: 10.3390/biom11050767. Biomolecules. 2021. PMID: 34065606 Free PMC article. Review.
Cited by
-
Low dose DMSO treatment induces oligomerization and accelerates aggregation of α-synuclein.Sci Rep. 2022 Mar 8;12(1):3737. doi: 10.1038/s41598-022-07706-2. Sci Rep. 2022. PMID: 35260646 Free PMC article.
-
Uncovering the Role of Natural and Synthetic Small Molecules in Counteracting the Burden of α-Synuclein Aggregates and Related Toxicity in Different Models of Parkinson's Disease.Int J Mol Sci. 2023 Aug 29;24(17):13370. doi: 10.3390/ijms241713370. Int J Mol Sci. 2023. PMID: 37686175 Free PMC article. Review.
-
Characterization of Pairs of Toxic and Nontoxic Misfolded Protein Oligomers Elucidates the Structural Determinants of Oligomer Toxicity in Protein Misfolding Diseases.Acc Chem Res. 2023 Jun 20;56(12):1395-1405. doi: 10.1021/acs.accounts.3c00045. Epub 2023 Apr 18. Acc Chem Res. 2023. PMID: 37071750 Free PMC article.
-
Recombinant Full-Length TDP-43 Oligomers Retain Their Ability to Bind RNAs, Are Not Toxic, and Do Not Seed TDP-43 Aggregation in Vitro.ACS Chem Neurosci. 2024 Jan 3;15(1):193-204. doi: 10.1021/acschemneuro.3c00691. Epub 2023 Dec 20. ACS Chem Neurosci. 2024. PMID: 38116987 Free PMC article.
-
Diagnostic and therapeutic agents that target alpha-synuclein in Parkinson's disease.J Neurol. 2022 Nov;269(11):5762-5786. doi: 10.1007/s00415-022-11267-9. Epub 2022 Jul 13. J Neurol. 2022. PMID: 35831620 Free PMC article. Review.
References
-
- Spillantini M.G., Schmidt M.L., Lee V.M.Y., Trojanowski J.Q., Jakes R., Goedert M. α-Synuclein in Lewy bodies. Nature. 1997;388:839. - PubMed
-
- Uversky V.N., Li J., Fink A.L. Evidence for a partially folded intermediate in alpha-synuclein fibril formation. J. Biol. Chem. 2001;276:10737–10744. - PubMed
-
- Winner B., Jappelli R., Maji S.K., Desplats P.A., Boyer L., Aigner S., Hetzer C., Loher T., Vilar M., Campioni S., Tzitzilonis C., Soragni A., Jessberger S., Mira H., Consiglio A. In vivo demonstration that alpha-synuclein oligomers are toxic. Proc. Natl. Acad. Sci. U. S. A. 2011;108:4194–4199. - PMC - PubMed
-
- Lorenzen N., Lemminger L., Pedersen J.N., Nielsen S.B., Otzen D.E. The N-terminus of alpha-synuclein is essential for both monomeric and oligomeric interactions with membranes. FEBS Lett. 2014;588:497–502. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
