

AP-3-dependent targeting of flippase ATP8A1 to lamellar bodies suppresses activation of YAP in alveolar epithelial type 2 cells
- PMID: 33990468
- PMCID: PMC8157957
- DOI: 10.1073/pnas.2025208118
AP-3-dependent targeting of flippase ATP8A1 to lamellar bodies suppresses activation of YAP in alveolar epithelial type 2 cells
Abstract
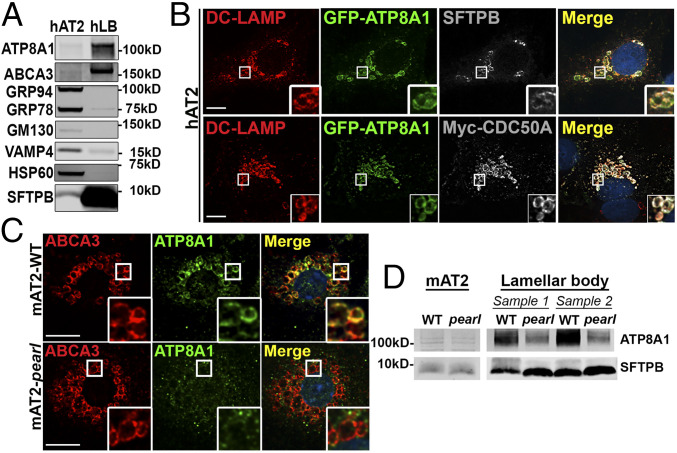
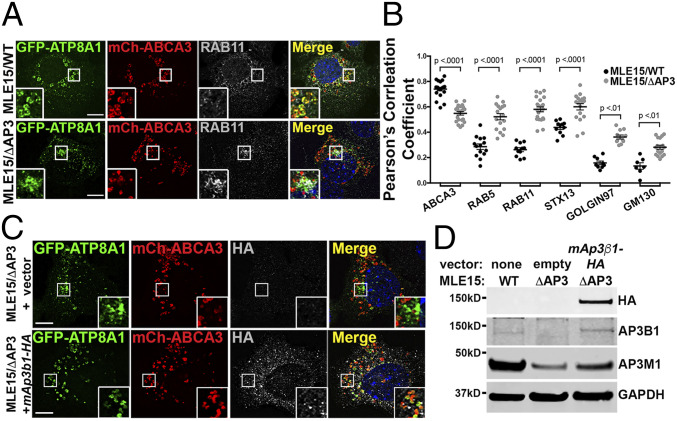
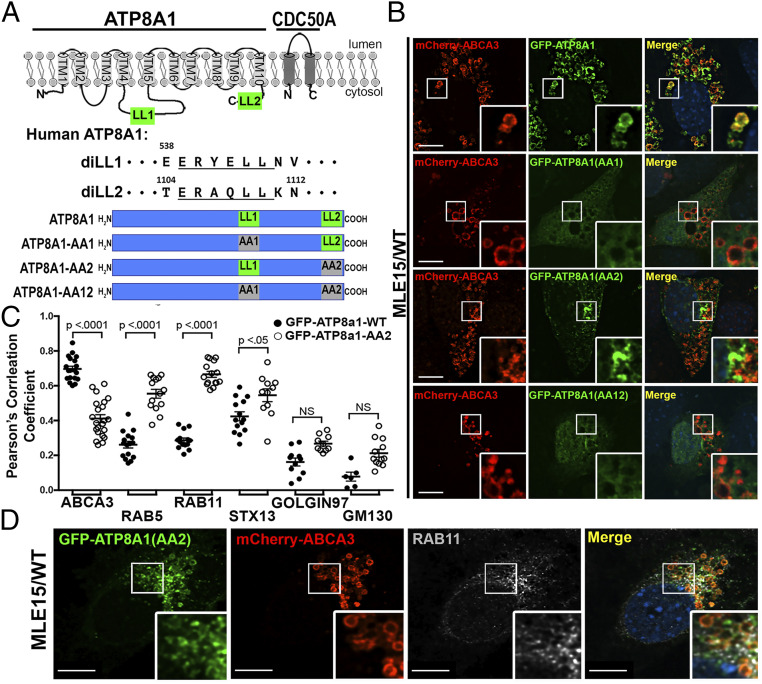
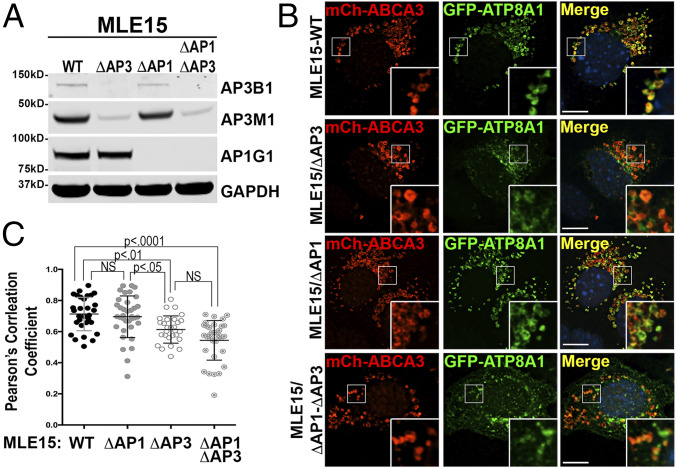
Lamellar bodies (LBs) are lysosome-related organelles (LROs) of surfactant-producing alveolar type 2 (AT2) cells of the distal lung epithelium. Trafficking pathways to LBs have been understudied but are likely critical to AT2 cell homeostasis given associations between genetic defects of endosome to LRO trafficking and pulmonary fibrosis in Hermansky Pudlak syndrome (HPS). Our prior studies uncovered a role for AP-3, defective in HPS type 2, in trafficking Peroxiredoxin-6 to LBs. We now show that the P4-type ATPase ATP8A1 is sorted by AP-3 from early endosomes to LBs through recognition of a C-terminal dileucine-based signal. Disruption of the AP-3/ATP8A1 interaction causes ATP8A1 accumulation in early sorting and/or recycling endosomes, enhancing phosphatidylserine exposure on the cytosolic leaflet. This in turn promotes activation of Yes-activating protein, a transcriptional coactivator, augmenting cell migration and AT2 cell numbers. Together, these studies illuminate a mechanism whereby loss of AP-3-mediated trafficking contributes to a toxic gain-of-function that results in enhanced and sustained activation of a repair pathway associated with pulmonary fibrosis.
Keywords: endosome; lung epithelium; lysosome; pulmonary fibrosis.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Impaired Lysosomal Integral Membrane Protein 2-dependent Peroxiredoxin 6 Delivery to Lamellar Bodies Accounts for Altered Alveolar Phospholipid Content in Adaptor Protein-3-deficient pearl Mice.J Biol Chem. 2016 Apr 15;291(16):8414-27. doi: 10.1074/jbc.M116.720201. Epub 2016 Feb 23. J Biol Chem. 2016. PMID: 26907692 Free PMC article.
-
Dysregulated alveolar epithelial cell progenitor function and identity in Hermansky-Pudlak syndrome.JCI Insight. 2024 Dec 19;10(3):e183483. doi: 10.1172/jci.insight.183483. JCI Insight. 2024. PMID: 39699958
-
Endosomal phosphatidylserine is critical for the YAP signalling pathway in proliferating cells.Nat Commun. 2017 Nov 1;8(1):1246. doi: 10.1038/s41467-017-01255-3. Nat Commun. 2017. PMID: 29093443 Free PMC article.
-
Pathogenesis and Therapy of Hermansky-Pudlak Syndrome (HPS)-Associated Pulmonary Fibrosis.Int J Mol Sci. 2024 Oct 19;25(20):11270. doi: 10.3390/ijms252011270. Int J Mol Sci. 2024. PMID: 39457053 Free PMC article. Review.
-
AP-3-dependent trafficking and disease: the first decade.Curr Opin Cell Biol. 2009 Aug;21(4):552-9. doi: 10.1016/j.ceb.2009.04.014. Epub 2009 Jun 3. Curr Opin Cell Biol. 2009. PMID: 19497727 Review.
Cited by
-
The Role of Hippo/YAP Signaling in Alveolar Repair and Pulmonary Fibrosis.Front Med (Lausanne). 2021 Oct 4;8:752316. doi: 10.3389/fmed.2021.752316. eCollection 2021. Front Med (Lausanne). 2021. PMID: 34671628 Free PMC article. Review.
-
UniTmp: unified resources for transmembrane proteins.Nucleic Acids Res. 2024 Jan 5;52(D1):D572-D578. doi: 10.1093/nar/gkad897. Nucleic Acids Res. 2024. PMID: 37870462 Free PMC article.
-
Alveolar Organoids in Lung Disease Modeling.Biomolecules. 2024 Jan 16;14(1):115. doi: 10.3390/biom14010115. Biomolecules. 2024. PMID: 38254715 Free PMC article. Review.
-
Insights into the Pathogenesis of Pulmonary Fibrosis from Genetic Diseases.Am J Respir Cell Mol Biol. 2022 Jul;67(1):20-35. doi: 10.1165/rcmb.2021-0557TR. Am J Respir Cell Mol Biol. 2022. PMID: 35294321 Free PMC article. Review.
-
Channels and Transporters of the Pulmonary Lamellar Body in Health and Disease.Cells. 2021 Dec 24;11(1):45. doi: 10.3390/cells11010045. Cells. 2021. PMID: 35011607 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
