

Lysosome biogenesis: Regulation and functions
- PMID: 33950241
- PMCID: PMC8105738
- DOI: 10.1083/jcb.202102001
Lysosome biogenesis: Regulation and functions
Abstract
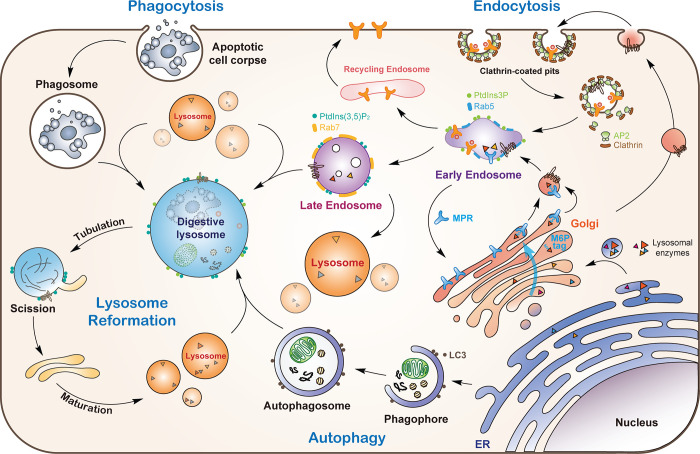
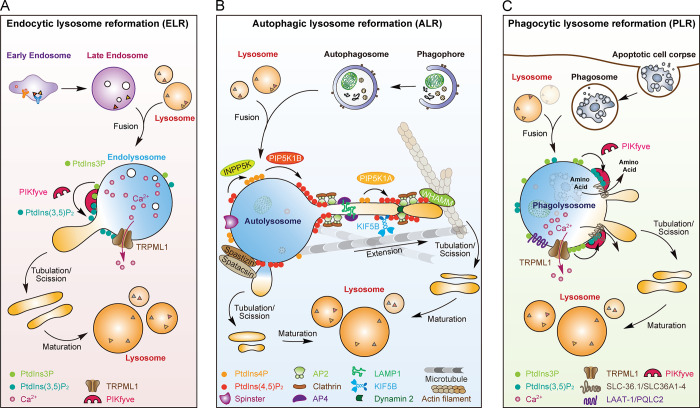
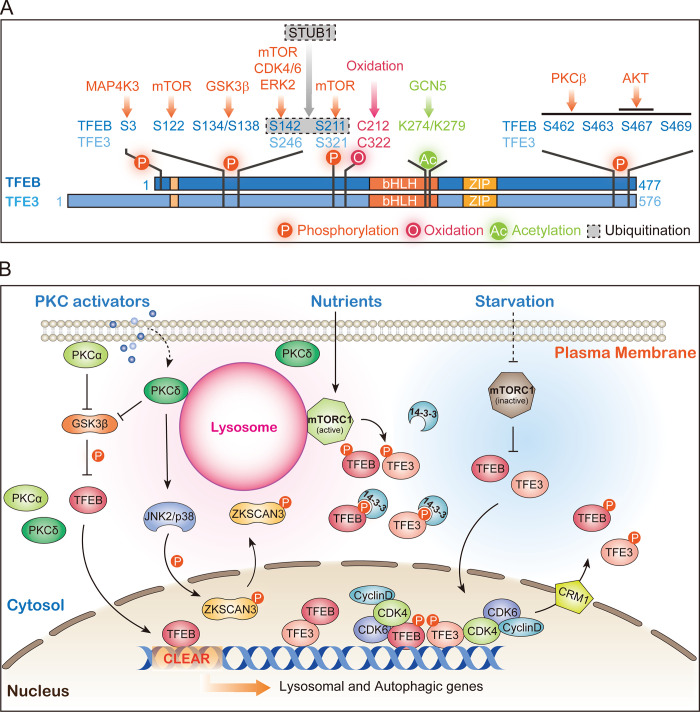
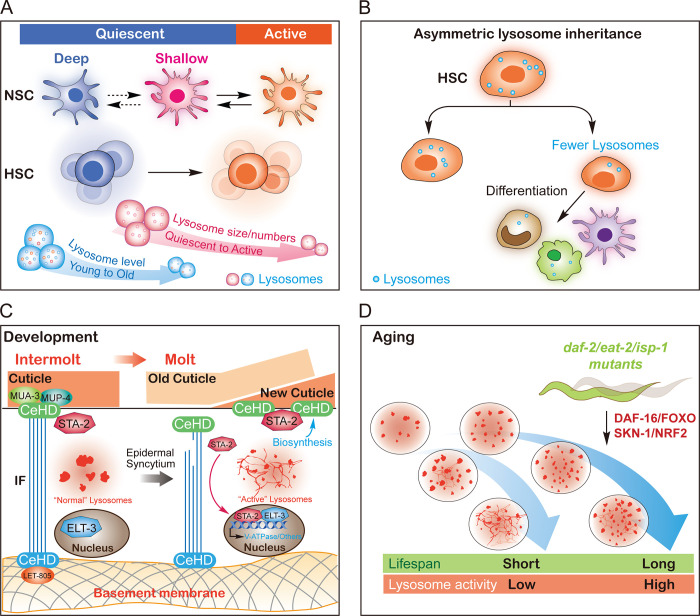
Lysosomes are degradation centers and signaling hubs in cells and play important roles in cellular homeostasis, development, and aging. Changes in lysosome function are essential to support cellular adaptation to multiple signals and stimuli. Therefore, lysosome biogenesis and activity are regulated by a wide variety of intra- and extracellular cues. Here, we summarize current knowledge of the regulatory mechanisms of lysosome biogenesis, including synthesis of lysosomal proteins and their delivery via the endosome-lysosome pathway, reformation of lysosomes from degradative vesicles, and transcriptional regulation of lysosomal genes. We survey the regulation of lysosome biogenesis in response to nutrient and nonnutrient signals, the cell cycle, stem cell quiescence, and cell fate determination. Finally, we discuss lysosome biogenesis and functions in the context of organismal development and aging.
© 2021 Yang and Wang.
Figures
Similar articles
-
Lysosomes at the Crossroads of Cell Metabolism, Cell Cycle, and Stemness.Int J Mol Sci. 2022 Feb 18;23(4):2290. doi: 10.3390/ijms23042290. Int J Mol Sci. 2022. PMID: 35216401 Free PMC article. Review.
-
Lysosomes as dynamic regulators of cell and organismal homeostasis.Nat Rev Mol Cell Biol. 2020 Feb;21(2):101-118. doi: 10.1038/s41580-019-0185-4. Epub 2019 Nov 25. Nat Rev Mol Cell Biol. 2020. PMID: 31768005 Review.
-
Molecular Mechanisms of Lysosome and Nucleus Communication.Trends Biochem Sci. 2020 Nov;45(11):978-991. doi: 10.1016/j.tibs.2020.06.004. Epub 2020 Jul 2. Trends Biochem Sci. 2020. PMID: 32624271 Free PMC article. Review.
-
Current methods to analyze lysosome morphology, positioning, motility and function.Traffic. 2022 May;23(5):238-269. doi: 10.1111/tra.12839. Epub 2022 Apr 24. Traffic. 2022. PMID: 35343629 Free PMC article. Review.
-
Lysosomal size matters.Traffic. 2020 Jan;21(1):60-75. doi: 10.1111/tra.12714. Epub 2019 Dec 6. Traffic. 2020. PMID: 31808235 Free PMC article. Review.
Cited by
-
The critical role of the endolysosomal system in cerebral ischemia.Neural Regen Res. 2023 May;18(5):983-990. doi: 10.4103/1673-5374.355745. Neural Regen Res. 2023. PMID: 36254978 Free PMC article. Review.
-
De novo lipogenesis fuels adipocyte autophagosome and lysosome membrane dynamics.Nat Commun. 2023 Mar 13;14(1):1362. doi: 10.1038/s41467-023-37016-8. Nat Commun. 2023. PMID: 36914626 Free PMC article.
-
Hypoxia promotes EV secretion by impairing lysosomal homeostasis in HNSCC through negative regulation of ATP6V1A by HIF-1α.J Extracell Vesicles. 2023 Feb;12(2):e12310. doi: 10.1002/jev2.12310. J Extracell Vesicles. 2023. PMID: 36748335 Free PMC article.
-
GSK3 inhibitor enhances gemtuzumab ozogamicin-induced apoptosis in primary human leukemia cells by overcoming multiple mechanisms of resistance.EJHaem. 2022 Dec 12;4(1):153-164. doi: 10.1002/jha2.600. eCollection 2023 Feb. EJHaem. 2022. PMID: 36819180 Free PMC article.
-
Age-Related Lysosomal Dysfunctions.Cells. 2022 Jun 20;11(12):1977. doi: 10.3390/cells11121977. Cells. 2022. PMID: 35741106 Free PMC article. Review.
References
-
- Annunziata, I., van de Vlekkert D., Wolf E., Finkelstein D., Neale G., Machado E., Mosca R., Campos Y., Tillman H., Roussel M.F., et al. . 2019. MYC competes with MiT/TFE in regulating lysosomal biogenesis and autophagy through an epigenetic rheostat. Nat. Commun. 10:3623. 10.1038/s41467-019-11568-0 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
