

A complex genetic architecture in zebrafish relatives Danio quagga and D. kyathit underlies development of stripes and spots
- PMID: 33901178
- PMCID: PMC8102007
- DOI: 10.1371/journal.pgen.1009364
A complex genetic architecture in zebrafish relatives Danio quagga and D. kyathit underlies development of stripes and spots
Abstract
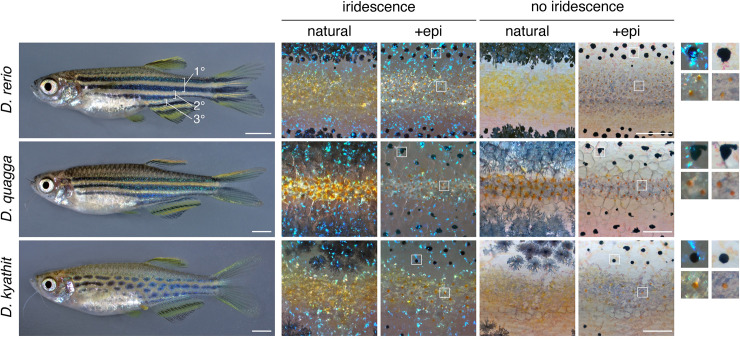
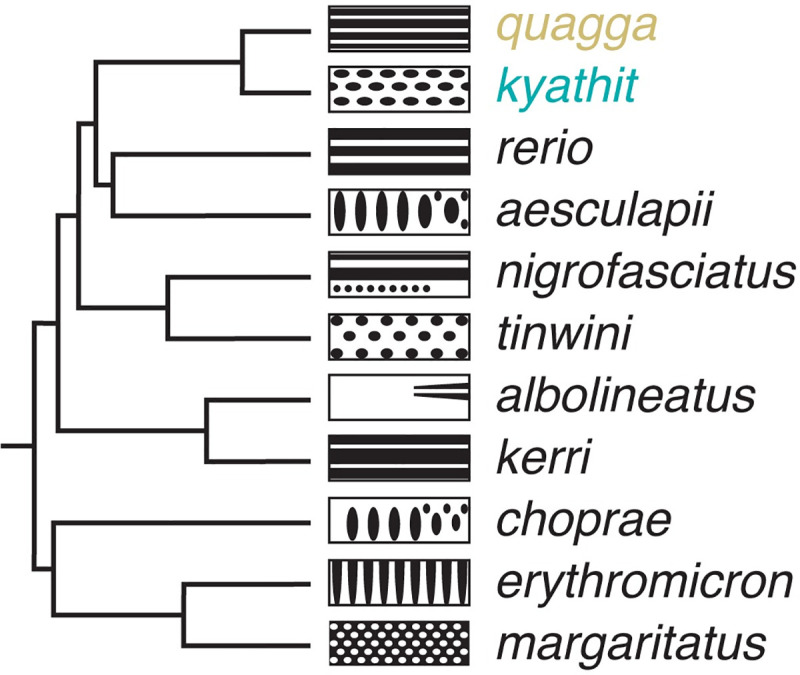
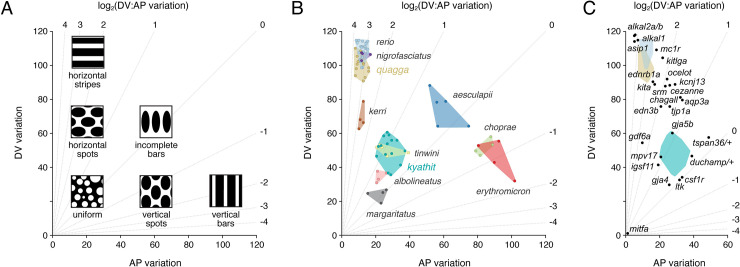
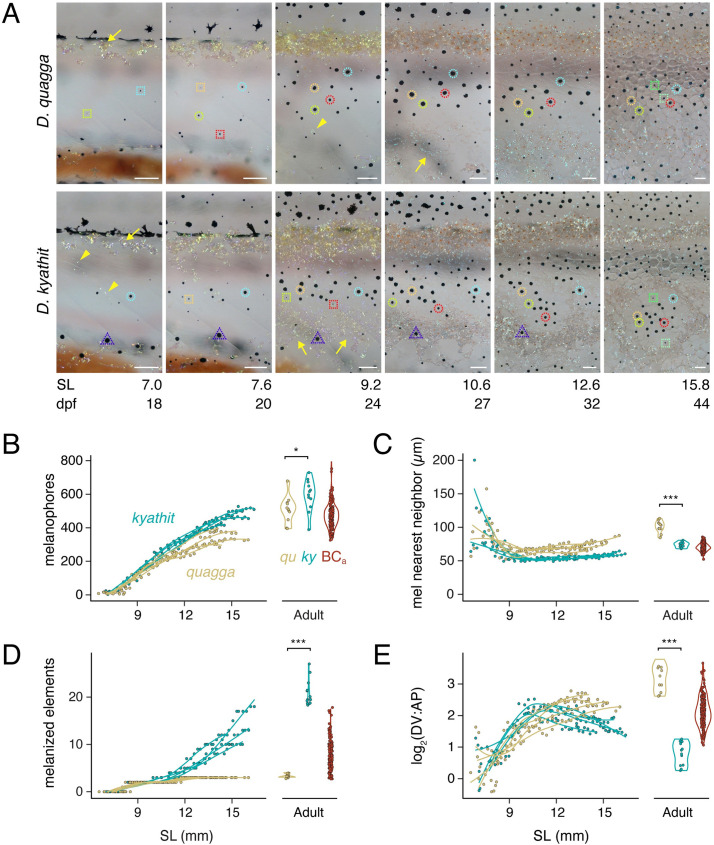
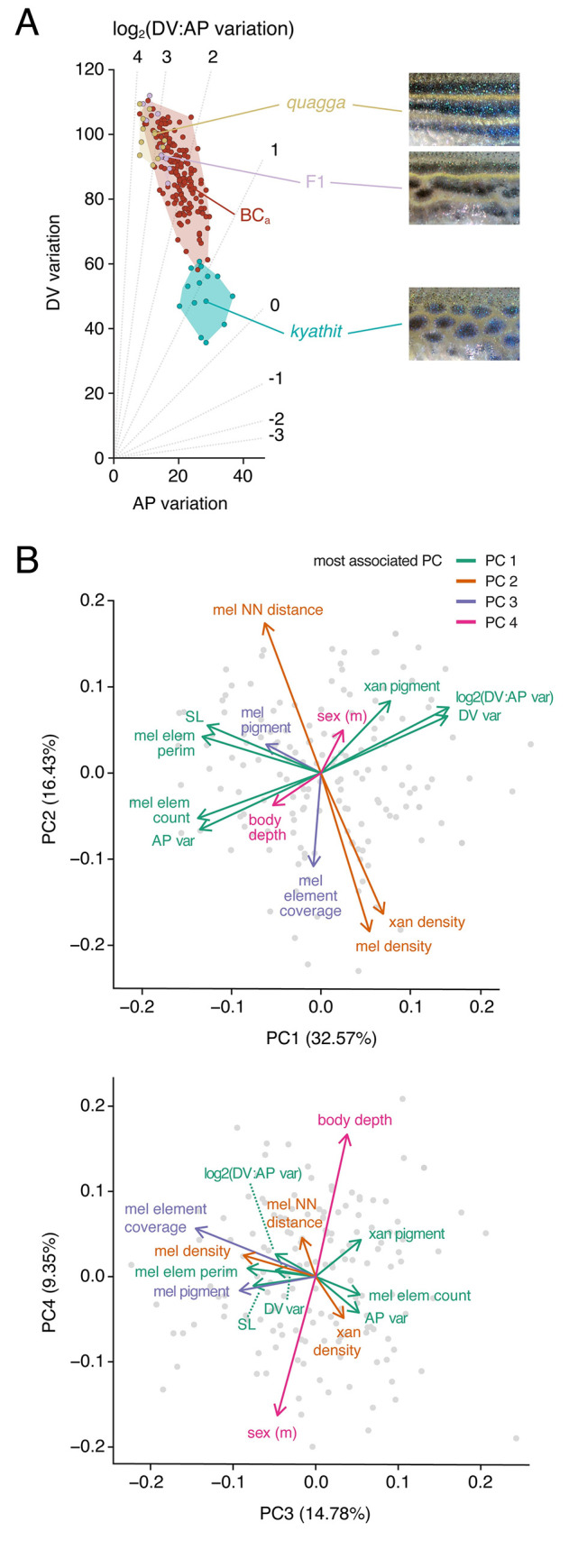
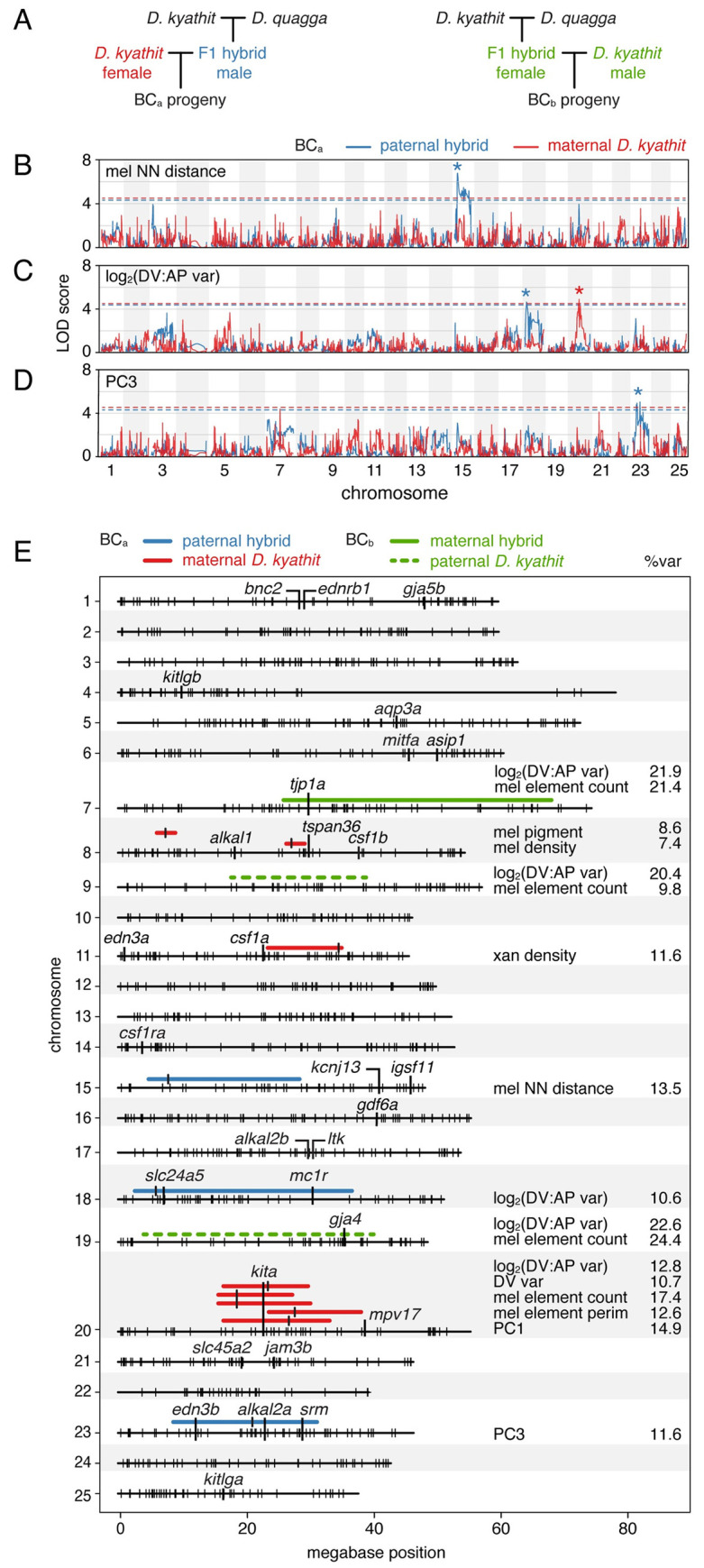
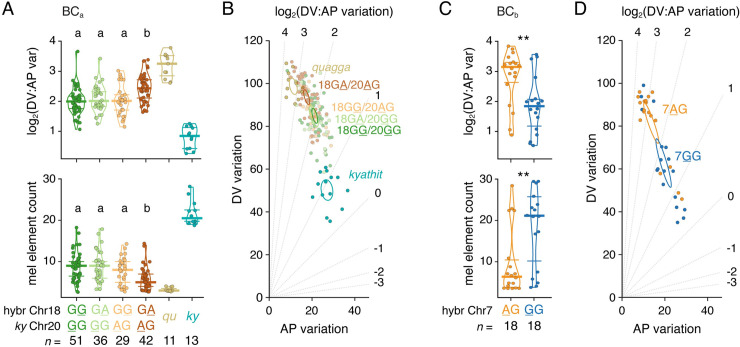
Vertebrate pigmentation is a fundamentally important, multifaceted phenotype. Zebrafish, Danio rerio, has been a valuable model for understanding genetics and development of pigment pattern formation due to its genetic and experimental tractability, advantages that are shared across several Danio species having a striking array of pigment patterns. Here, we use the sister species D. quagga and D. kyathit, with stripes and spots, respectively, to understand how natural genetic variation impacts phenotypes at cellular and organismal levels. We first show that D. quagga and D. kyathit phenotypes resemble those of wild-type D. rerio and several single locus mutants of D. rerio, respectively, in a morphospace defined by pattern variation along dorsoventral and anteroposterior axes. We then identify differences in patterning at the cellular level between D. quagga and D. kyathit by repeated daily imaging during pattern development and quantitative comparisons of adult phenotypes, revealing that patterns are similar initially but diverge ontogenetically. To assess the genetic architecture of these differences, we employ reduced-representation sequencing of second-generation hybrids. Despite the similarity of D. quagga to D. rerio, and D. kyathit to some D. rerio mutants, our analyses reveal a complex genetic basis for differences between D. quagga and D. kyathit, with several quantitative trait loci contributing to variation in overall pattern and cellular phenotypes, epistatic interactions between loci, and abundant segregating variation within species. Our findings provide a window into the evolutionary genetics of pattern-forming mechanisms in Danio and highlight the complexity of differences that can arise even between sister species. Further studies of natural genetic diversity underlying pattern variation in D. quagga and D. kyathit should provide insights complementary to those from zebrafish mutant phenotypes and more distant species comparisons.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Zebrafish hybrids suggest genetic mechanisms for pigment pattern diversification in Danio.Dev Genes Evol. 2001 Jul;211(7):319-28. doi: 10.1007/s004270100155. Dev Genes Evol. 2001. PMID: 11466528
-
Evolutionary diversification of pigment pattern in Danio fishes: differential fms dependence and stripe loss in D. albolineatus.Development. 2005 Jan;132(1):89-104. doi: 10.1242/dev.01547. Epub 2004 Nov 24. Development. 2005. PMID: 15563521
-
Deconstructing evolution of adult phenotypes: genetic analyses of kit reveal homology and evolutionary novelty during adult pigment pattern development of Danio fishes.Development. 2007 Mar;134(6):1081-90. doi: 10.1242/dev.02799. Epub 2007 Feb 7. Development. 2007. PMID: 17287252
-
The Developmental Genetics of Vertebrate Color Pattern Formation: Lessons from Zebrafish.Curr Top Dev Biol. 2016;117:141-69. doi: 10.1016/bs.ctdb.2015.12.012. Epub 2016 Feb 23. Curr Top Dev Biol. 2016. PMID: 26969976 Review.
-
Zebrafish stripes as a model for vertebrate colour pattern formation.Curr Biol. 2015 Jan 19;25(2):R81-R92. doi: 10.1016/j.cub.2014.11.013. Curr Biol. 2015. PMID: 25602311 Review.
Cited by
-
kcnj13 regulates pigment cell shapes in zebrafish and has diverged by cis-regulatory evolution between Danio species.Development. 2023 Aug 15;150(16):dev201627. doi: 10.1242/dev.201627. Epub 2023 Aug 24. Development. 2023. PMID: 37530080 Free PMC article.
-
Pigment pattern morphospace of Danio fishes: evolutionary diversification and mutational effects.Biol Open. 2021 Sep 15;10(9):bio058814. doi: 10.1242/bio.058814. Epub 2021 Sep 20. Biol Open. 2021. PMID: 34463758 Free PMC article.
-
Development and genetics of red coloration in the zebrafish relative Danio albolineatus.Elife. 2021 Aug 26;10:e70253. doi: 10.7554/eLife.70253. Elife. 2021. PMID: 34435950 Free PMC article.
-
A highly contiguous nuclear genome assembly of the mandarinfish Synchiropus splendidus (Syngnathiformes: Callionymidae).G3 (Bethesda). 2021 Dec 8;11(12):jkab306. doi: 10.1093/g3journal/jkab306. G3 (Bethesda). 2021. PMID: 34849773 Free PMC article.
References
-
- Hamilton F. An Account of the Fishes found in the River Ganges and its Branches. Edinburgh: Archibald Constable and Company; 1822.
-
- Cartner SC, Eisen JS, Farmer SC, Guillemin KJ, Kent ML, Sanders GE. The Zebrafish in Biomedical Research: Academic Press; 2020. 2020/01/01/.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
