

Orchestral manoeuvres in the light: crosstalk needed for regulation of the Chlamydomonas carbon concentration mechanism
- PMID: 33893473
- PMCID: PMC8320531
- DOI: 10.1093/jxb/erab169
Orchestral manoeuvres in the light: crosstalk needed for regulation of the Chlamydomonas carbon concentration mechanism
Erratum in
-
Correction to: Orchestral manoeuvres in the light: crosstalk needed for regulation of the Chlamydomonas carbon concentration mechanism.J Exp Bot. 2022 Aug 11;73(14):5084. doi: 10.1093/jxb/erac272. J Exp Bot. 2022. PMID: 35861227 Free PMC article. No abstract available.
Abstract
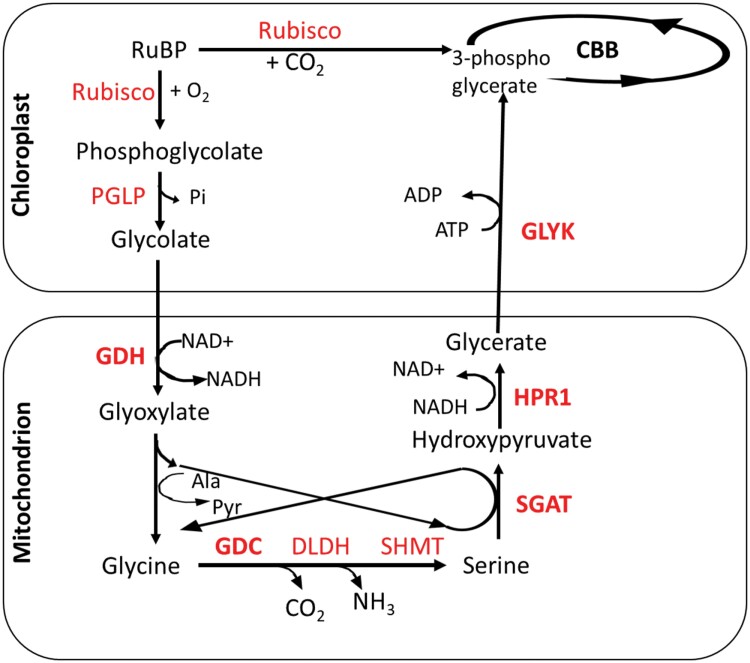
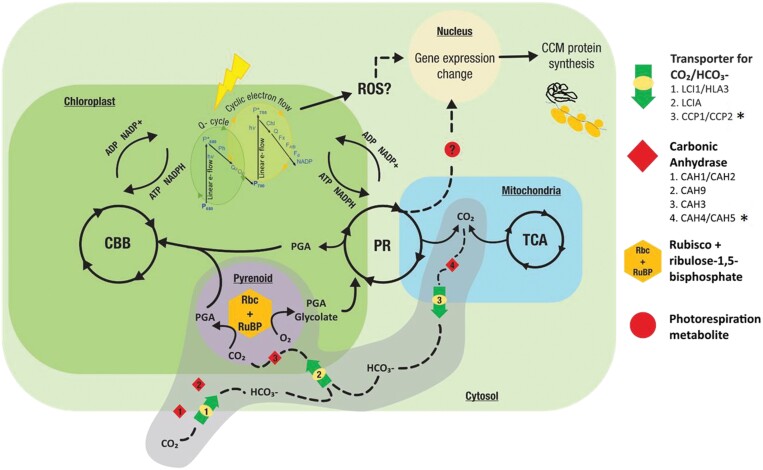
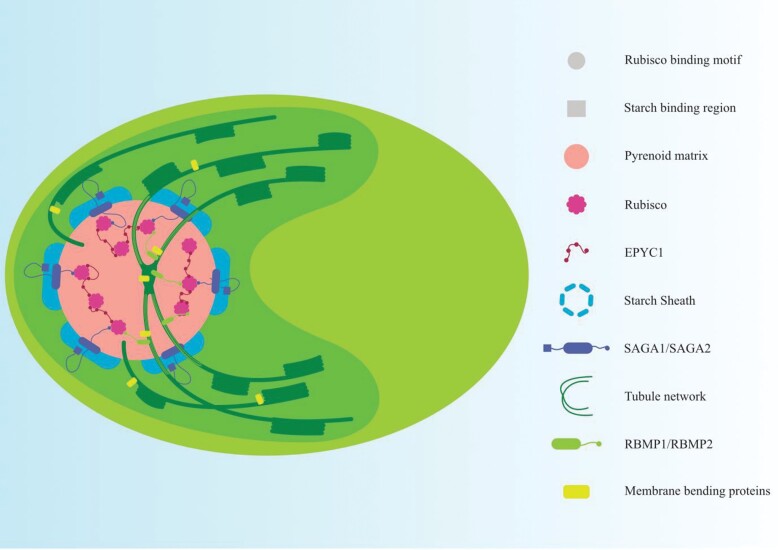
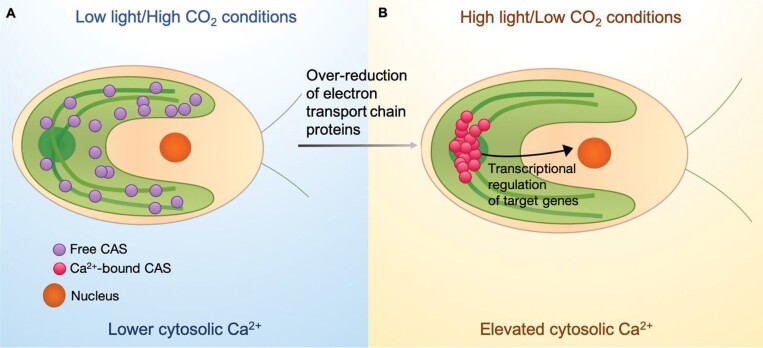
The inducible carbon concentration mechanism (CCM) in Chlamydomonas reinhardtii has been well defined from a molecular and ultrastructural perspective. Inorganic carbon transport proteins, and strategically located carbonic anhydrases deliver CO2 within the chloroplast pyrenoid matrix where Rubisco is packaged. However, there is little understanding of the fundamental signalling and sensing processes leading to CCM induction. While external CO2 limitation has been believed to be the primary cue, the coupling between energetic supply and inorganic carbon demand through regulatory feedback from light harvesting and photorespiration signals could provide the original CCM trigger. Key questions regarding the integration of these processes are addressed in this review. We consider how the chloroplast functions as a crucible for photosynthesis, importing and integrating nuclear-encoded components from the cytoplasm, and sending retrograde signals to the nucleus to regulate CCM induction. We hypothesize that induction of the CCM is associated with retrograde signals associated with photorespiration and/or light stress. We have also examined the significance of common evolutionary pressures for origins of two co-regulated processes, namely the CCM and photorespiration, in addition to identifying genes of interest involved in transcription, protein folding, and regulatory processes which are needed to fully understand the processes leading to CCM induction.
Keywords: Chlamydomonas; CIA5; Carbon concentration mechanism (CCM); chaperones; photorespiration; photosynthesis; pyrenoid; retrograde signalling.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures

Similar articles
-
Mitochondrial carbonic anhydrases are needed for optimal photosynthesis at low CO2 levels in Chlamydomonas.Plant Physiol. 2021 Nov 3;187(3):1387-1398. doi: 10.1093/plphys/kiab351. Plant Physiol. 2021. PMID: 34618049 Free PMC article.
-
Light and low-CO2-dependent LCIB-LCIC complex localization in the chloroplast supports the carbon-concentrating mechanism in Chlamydomonas reinhardtii.Plant Cell Physiol. 2010 Sep;51(9):1453-68. doi: 10.1093/pcp/pcq105. Epub 2010 Jul 21. Plant Cell Physiol. 2010. PMID: 20660228
-
Chloroplast-mediated regulation of CO2-concentrating mechanism by Ca2+-binding protein CAS in the green alga Chlamydomonas reinhardtii.Proc Natl Acad Sci U S A. 2016 Nov 1;113(44):12586-12591. doi: 10.1073/pnas.1606519113. Epub 2016 Oct 17. Proc Natl Acad Sci U S A. 2016. PMID: 27791081 Free PMC article.
-
The CO2 concentrating mechanism and photosynthetic carbon assimilation in limiting CO2 : how Chlamydomonas works against the gradient.Plant J. 2015 May;82(3):429-448. doi: 10.1111/tpj.12829. Plant J. 2015. PMID: 25765072 Review.
-
New horizons for building pyrenoid-based CO2-concentrating mechanisms in plants to improve yields.Plant Physiol. 2022 Oct 27;190(3):1609-1627. doi: 10.1093/plphys/kiac373. Plant Physiol. 2022. PMID: 35961043 Free PMC article. Review.
Cited by
-
Carbonic anhydrase versatility: from pH regulation to CO2 sensing and metabolism.Front Mol Biosci. 2023 Nov 10;10:1326633. doi: 10.3389/fmolb.2023.1326633. eCollection 2023. Front Mol Biosci. 2023. PMID: 38028557 Free PMC article.
-
Crossing and selection of Chlamydomonas reinhardtii strains for biotechnological glycolate production.Appl Microbiol Biotechnol. 2022 May;106(9-10):3539-3554. doi: 10.1007/s00253-022-11933-y. Epub 2022 May 5. Appl Microbiol Biotechnol. 2022. PMID: 35511277 Free PMC article.
-
Adapting from Low to High: An Update to CO2-Concentrating Mechanisms of Cyanobacteria and Microalgae.Plants (Basel). 2023 Apr 6;12(7):1569. doi: 10.3390/plants12071569. Plants (Basel). 2023. PMID: 37050194 Free PMC article. Review.
-
The pyrenoid: the eukaryotic CO2-concentrating organelle.Plant Cell. 2023 Sep 1;35(9):3236-3259. doi: 10.1093/plcell/koad157. Plant Cell. 2023. PMID: 37279536 Free PMC article.
-
Regulation of Microalgal Photosynthetic Electron Transfer.Plants (Basel). 2024 Jul 29;13(15):2103. doi: 10.3390/plants13152103. Plants (Basel). 2024. PMID: 39124221 Free PMC article. Review.
References
-
- Arias C, Obudulub O, Zhaoa X, et al. 2020. Nuclear proteome analysis of Chlamydomonas with response to CO2 limitation. Algal Research 46, 101765.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
