

Classical MHC expression by DP thymocytes impairs the selection of non-classical MHC restricted innate-like T cells
- PMID: 33863906
- PMCID: PMC8052364
- DOI: 10.1038/s41467-021-22589-z
Classical MHC expression by DP thymocytes impairs the selection of non-classical MHC restricted innate-like T cells
Abstract
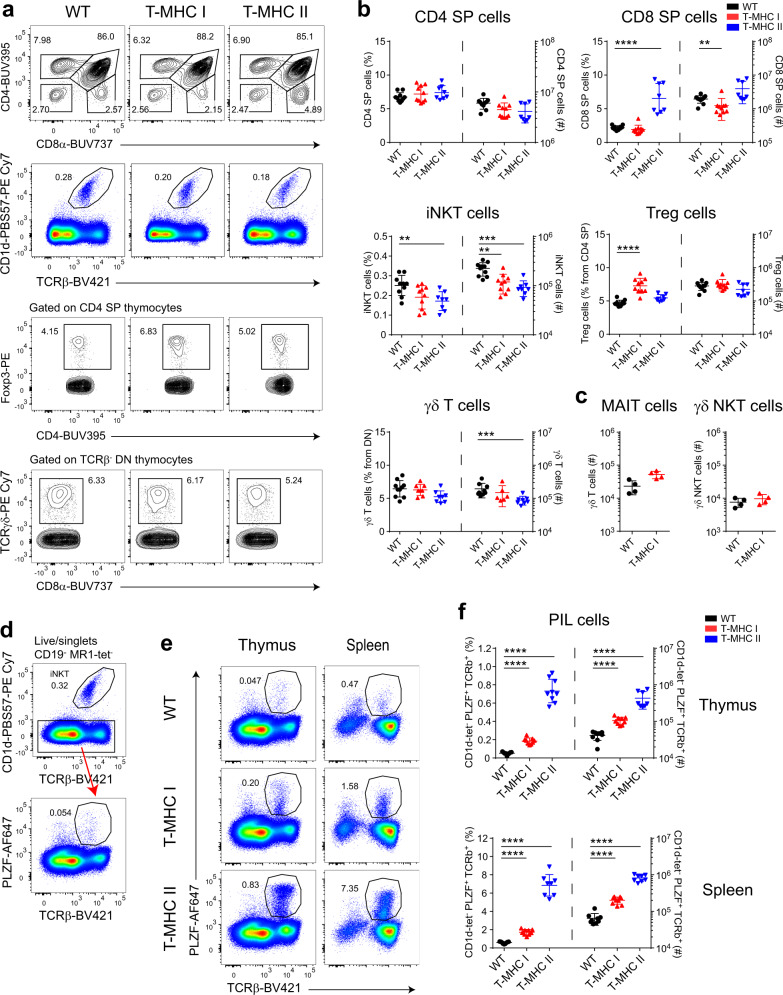
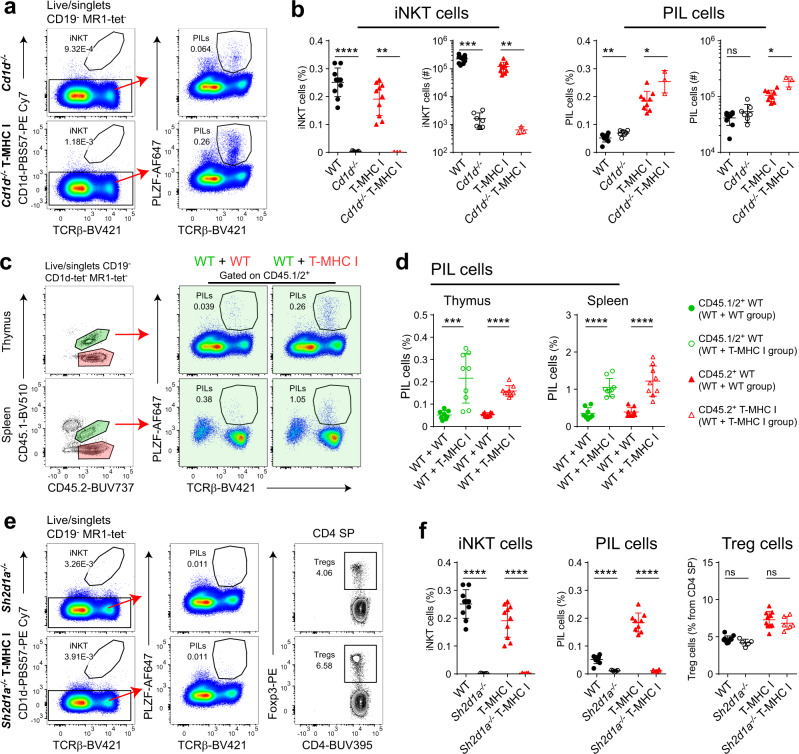
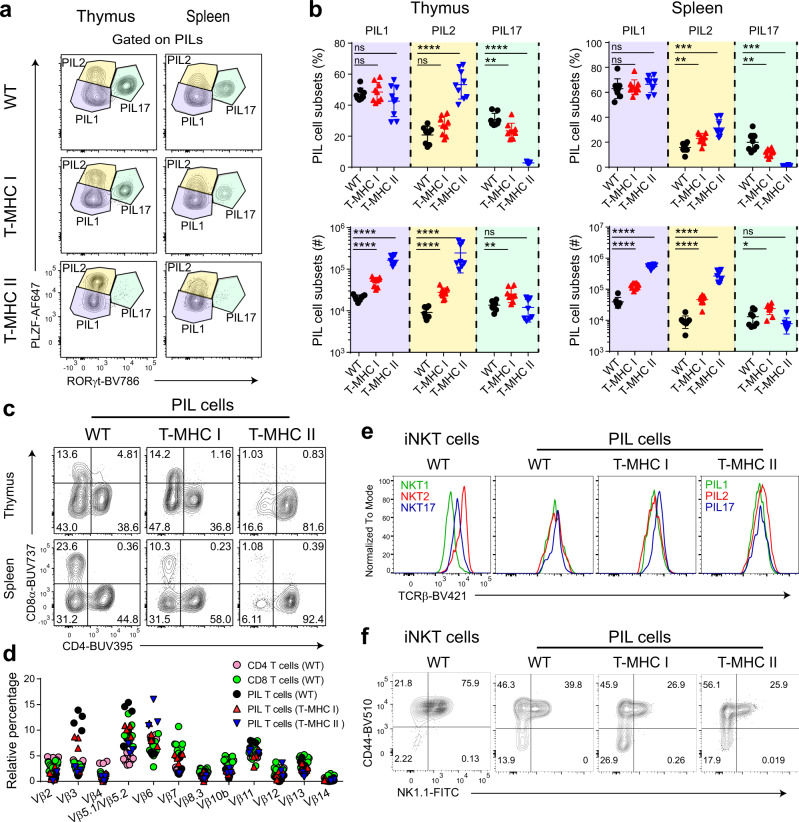
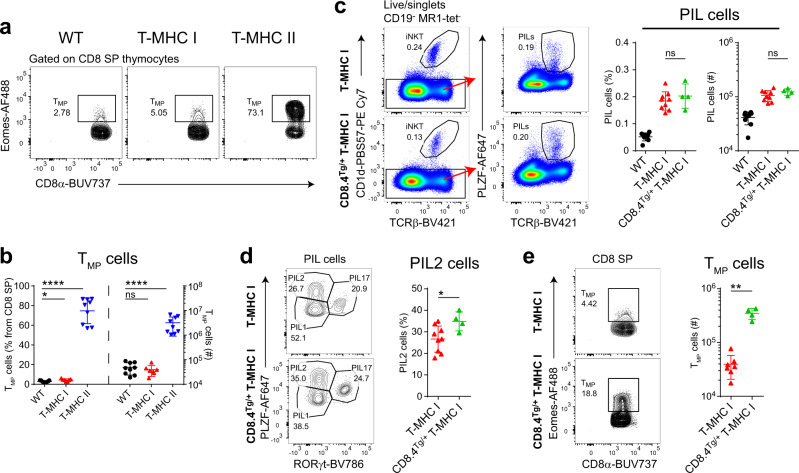
Conventional T cells are selected by peptide-MHC expressed by cortical epithelial cells in the thymus, and not by cortical thymocytes themselves that do not express MHC I or MHC II. Instead, cortical thymocytes express non-peptide presenting MHC molecules like CD1d and MR1, and promote the selection of PLZF+ iNKT and MAIT cells, respectively. Here, we report an inducible class-I transactivator mouse that enables the expression of peptide presenting MHC I molecules in different cell types. We show that MHC I expression in DP thymocytes leads to expansion of peptide specific PLZF+ innate-like (PIL) T cells. Akin to iNKT cells, PIL T cells differentiate into three functional effector subsets in the thymus, and are dependent on SAP signaling. We demonstrate that PIL and NKT cells compete for a narrow niche, suggesting that the absence of peptide-MHC on DP thymocytes facilitates selection of non-peptide specific lymphocytes.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Stepwise development of MAIT cells in mouse and human.PLoS Biol. 2009 Mar 10;7(3):e54. doi: 10.1371/journal.pbio.1000054. PLoS Biol. 2009. PMID: 19278296 Free PMC article.
-
Innate PLZF+CD4+ αβ T cells develop and expand in the absence of Itk.J Immunol. 2014 Jul 15;193(2):673-87. doi: 10.4049/jimmunol.1302058. Epub 2014 Jun 13. J Immunol. 2014. PMID: 24928994 Free PMC article.
-
Molecular mechanisms of lineage decisions in metabolite-specific T cells.Nat Immunol. 2019 Sep;20(9):1244-1255. doi: 10.1038/s41590-019-0465-3. Epub 2019 Aug 20. Nat Immunol. 2019. PMID: 31431722
-
Role of CD1d- and MR1-Restricted T Cells in Asthma.Front Immunol. 2018 Aug 28;9:1942. doi: 10.3389/fimmu.2018.01942. eCollection 2018. Front Immunol. 2018. PMID: 30210497 Free PMC article. Review.
-
MAIT cell development in mice and humans.Mol Immunol. 2021 Feb;130:31-36. doi: 10.1016/j.molimm.2020.12.003. Epub 2020 Dec 19. Mol Immunol. 2021. PMID: 33352411 Review.
Cited by
-
The Role of Zinc in the T-Cell Metabolism in Infection Requires Further Investigation - An Opinion.Front Immunol. 2022 Mar 10;13:865504. doi: 10.3389/fimmu.2022.865504. eCollection 2022. Front Immunol. 2022. PMID: 35359968 Free PMC article. No abstract available.
-
Mapping the developing human immune system across organs.Science. 2022 Jun 3;376(6597):eabo0510. doi: 10.1126/science.abo0510. Epub 2022 Jun 3. Science. 2022. PMID: 35549310 Free PMC article.
-
Developmental trajectory of unconventional T cells of the cynomolgus macaque thymus.Heliyon. 2024 Oct 23;10(21):e39736. doi: 10.1016/j.heliyon.2024.e39736. eCollection 2024 Nov 15. Heliyon. 2024. PMID: 39524802 Free PMC article.
-
CD1b glycoprotein, a crucial marker of thymocyte development during T cell maturation in cynomolgus monkeys.Sci Rep. 2023 Sep 1;13(1):14388. doi: 10.1038/s41598-023-41708-y. Sci Rep. 2023. PMID: 37658106 Free PMC article.
-
MAIT cells: Conserved watchers on the wall.J Exp Med. 2025 Jan 6;222(1):e20232298. doi: 10.1084/jem.20232298. Epub 2024 Oct 24. J Exp Med. 2025. PMID: 39446132 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
