

Ago2 protects Drosophila siRNAs and microRNAs from target-directed degradation, even in the absence of 2'- O-methylation
- PMID: 33853897
- PMCID: PMC8127995
- DOI: 10.1261/rna.078746.121
Ago2 protects Drosophila siRNAs and microRNAs from target-directed degradation, even in the absence of 2'- O-methylation
Erratum in
-
Corrigendum: Ago2 protects Drosophila siRNAs and microRNAs from target-directed degradation, even in the absence of 2'-O-methylation.RNA. 2021 Dec;27(12):1617. doi: 10.1261/rna.078961.121. RNA. 2021. PMID: 34785574 Free PMC article. No abstract available.
Abstract
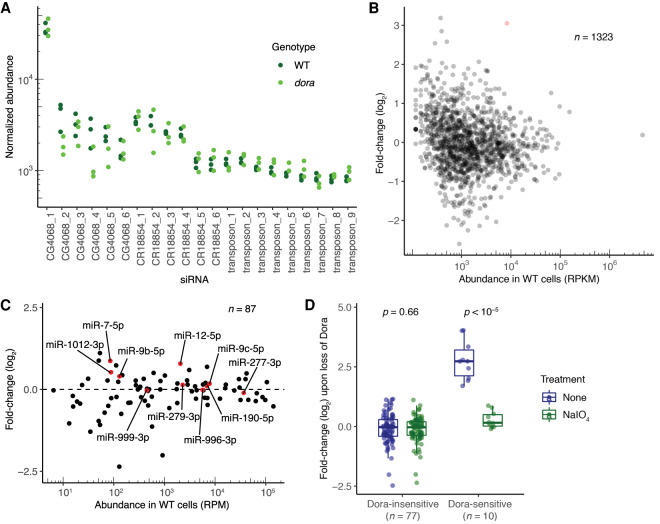
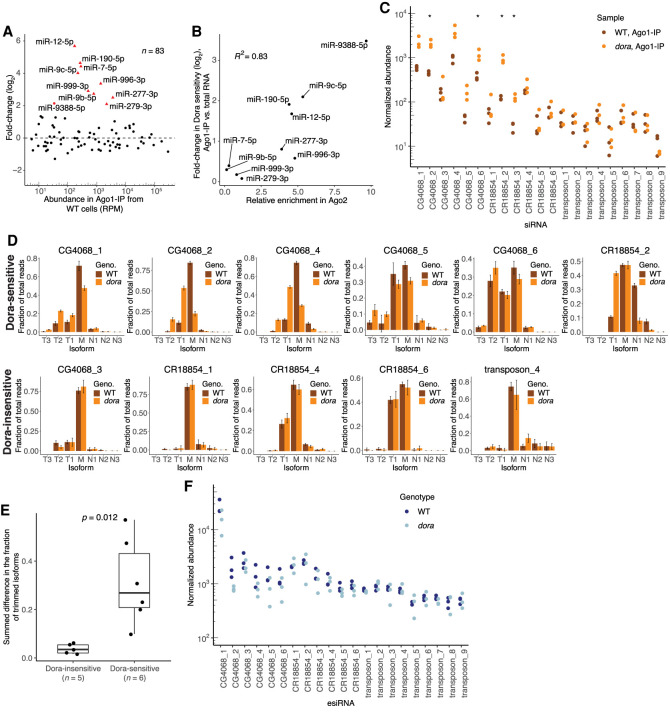
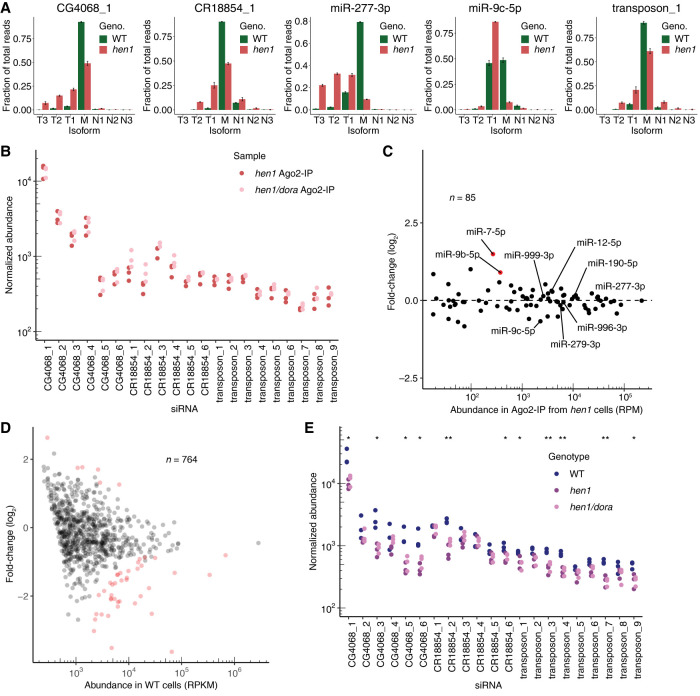
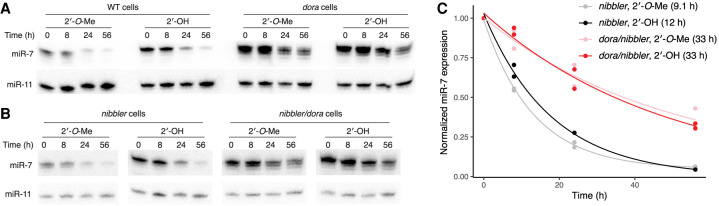
Target-directed microRNA (miRNA) degradation (TDMD), which is mediated by the protein ZSWIM8, plays a widespread role in shaping miRNA abundances across bilateria. Some endogenous small interfering RNAs (siRNAs) of Drosophila cells have target sites resembling those that trigger TDMD, raising the question as to whether they too might undergo such regulation by Dora, the Drosophila ZSWIM8 homolog. Here, we find that some of these siRNAs are indeed sensitive to Dora when loaded into Ago1, the Argonaute paralog that preferentially associates with miRNAs. Despite this sensitivity when loaded into Ago1, these siRNAs are not detectably regulated by target-directed degradation because most molecules are loaded into Ago2, the Argonaute paralog that preferentially associates with siRNAs, and we find that siRNAs and miRNAs loaded into Ago2 are insensitive to Dora. One explanation for the protection of these small RNAs loaded into Ago2 is that these small RNAs are 2'-O-methylated at their 3' termini. However, 2'-O-methylation does not protect these RNAs from Dora-mediated target-directed degradation, which indicates that their protection is instead conferred by features of the Ago2 protein itself. Together, these observations clarify the requirements for regulation by target-directed degradation and expand our understanding of the role of 2'-O-methylation in small-RNA biology.
Keywords: RNA methylation; TDMD; endogenous siRNAs; posttranscriptional regulation; siRNA dynamics; target-directed degradation.
© 2021 Kingston and Bartel; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
To kill a microRNA: emerging concepts in target-directed microRNA degradation.Nucleic Acids Res. 2024 Feb 28;52(4):1558-1574. doi: 10.1093/nar/gkae003. Nucleic Acids Res. 2024. PMID: 38224449 Free PMC article. Review.
-
The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation.Science. 2020 Dec 18;370(6523):eabc9359. doi: 10.1126/science.abc9359. Epub 2020 Nov 12. Science. 2020. PMID: 33184237 Free PMC article.
-
Sorting of Drosophila small silencing RNAs.Cell. 2007 Jul 27;130(2):299-308. doi: 10.1016/j.cell.2007.05.057. Cell. 2007. PMID: 17662944 Free PMC article.
-
R2D2 organizes small regulatory RNA pathways in Drosophila.Mol Cell Biol. 2011 Feb;31(4):884-96. doi: 10.1128/MCB.01141-10. Epub 2010 Dec 6. Mol Cell Biol. 2011. PMID: 21135122 Free PMC article.
-
Target-directed microRNA degradation: Mechanisms, significance, and functional implications.Wiley Interdiscip Rev RNA. 2024 Mar-Apr;15(2):e1832. doi: 10.1002/wrna.1832. Wiley Interdiscip Rev RNA. 2024. PMID: 38448799 Free PMC article. Review.
Cited by
-
To kill a microRNA: emerging concepts in target-directed microRNA degradation.Nucleic Acids Res. 2024 Feb 28;52(4):1558-1574. doi: 10.1093/nar/gkae003. Nucleic Acids Res. 2024. PMID: 38224449 Free PMC article. Review.
-
Endogenous transcripts direct microRNA degradation in Drosophila, and this targeted degradation is required for proper embryonic development.Mol Cell. 2022 Oct 20;82(20):3872-3884.e9. doi: 10.1016/j.molcel.2022.08.029. Epub 2022 Sep 22. Mol Cell. 2022. PMID: 36150386 Free PMC article.
-
Screening of Drosophila microRNA-degradation sequences reveals Argonaute1 mRNA's role in regulating miR-999.Nat Commun. 2023 Apr 13;14(1):2108. doi: 10.1038/s41467-023-37819-9. Nat Commun. 2023. PMID: 37055443 Free PMC article.
-
MicroRNA turnover: a tale of tailing, trimming, and targets.Trends Biochem Sci. 2023 Jan;48(1):26-39. doi: 10.1016/j.tibs.2022.06.005. Epub 2022 Jul 7. Trends Biochem Sci. 2023. PMID: 35811249 Free PMC article. Review.
-
RNA-Based Therapy for Cryptosporidium parvum Infection: Proof-of-Concept Studies.Infect Immun. 2022 Jul 21;90(7):e0019622. doi: 10.1128/iai.00196-22. Epub 2022 Jun 1. Infect Immun. 2022. PMID: 35647663 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials