

Next-generation inward rectifier potassium channel modulators: discovery and molecular pharmacology
- PMID: 33826405
- PMCID: PMC8285633
- DOI: 10.1152/ajpcell.00548.2020
Next-generation inward rectifier potassium channel modulators: discovery and molecular pharmacology
Abstract
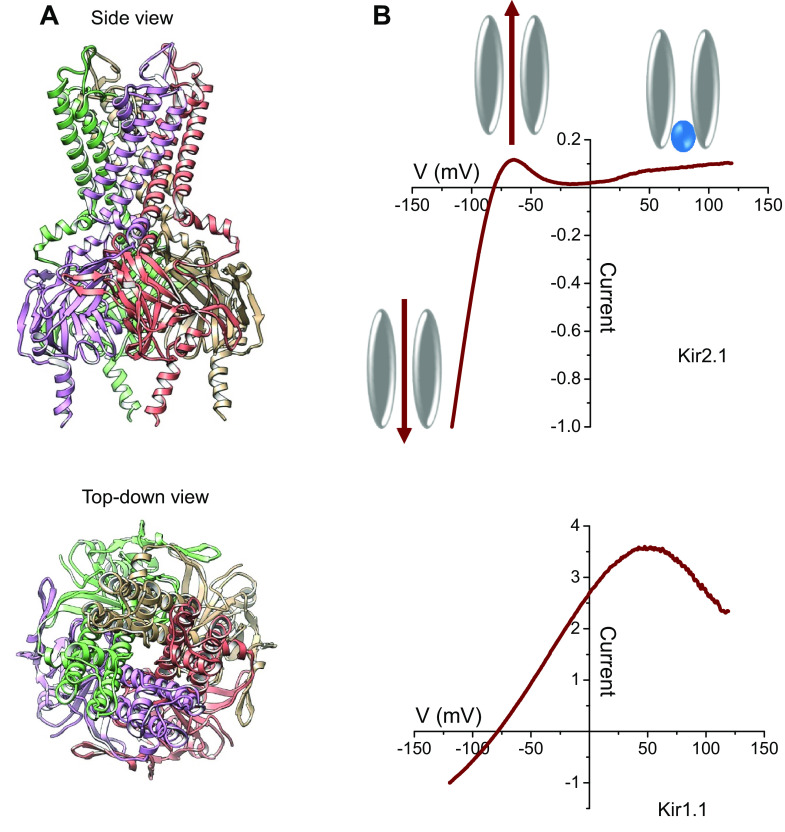
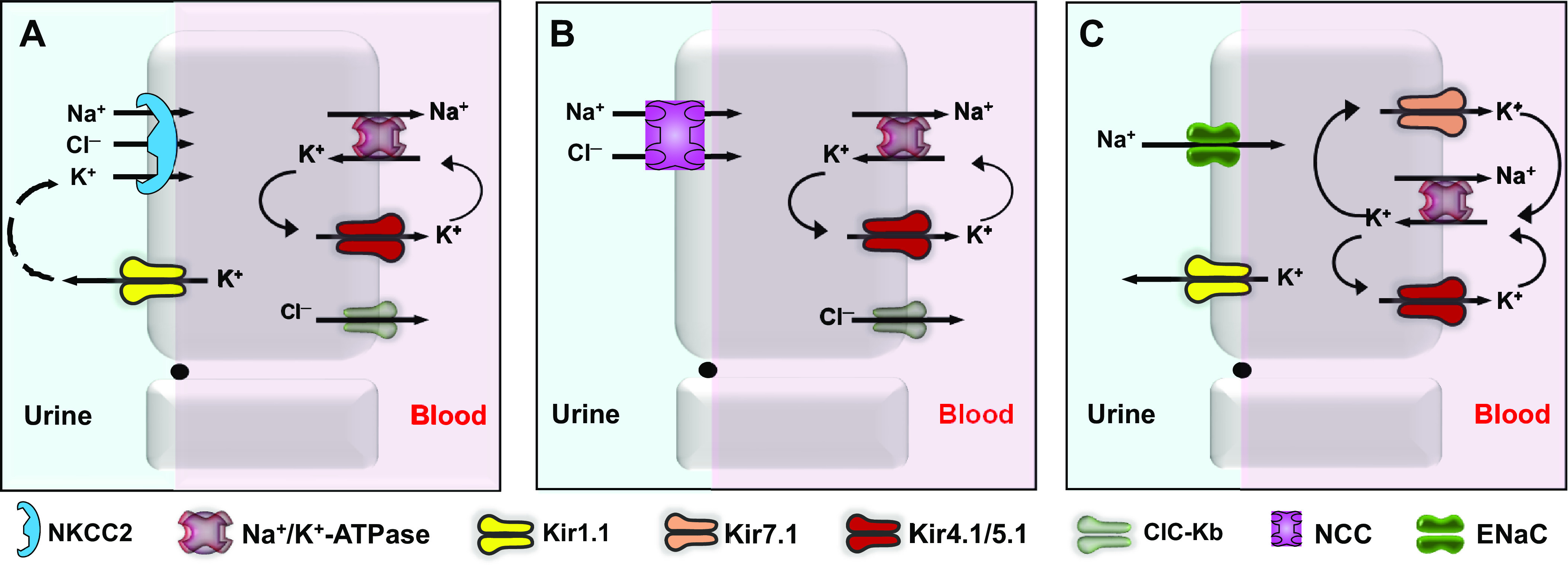
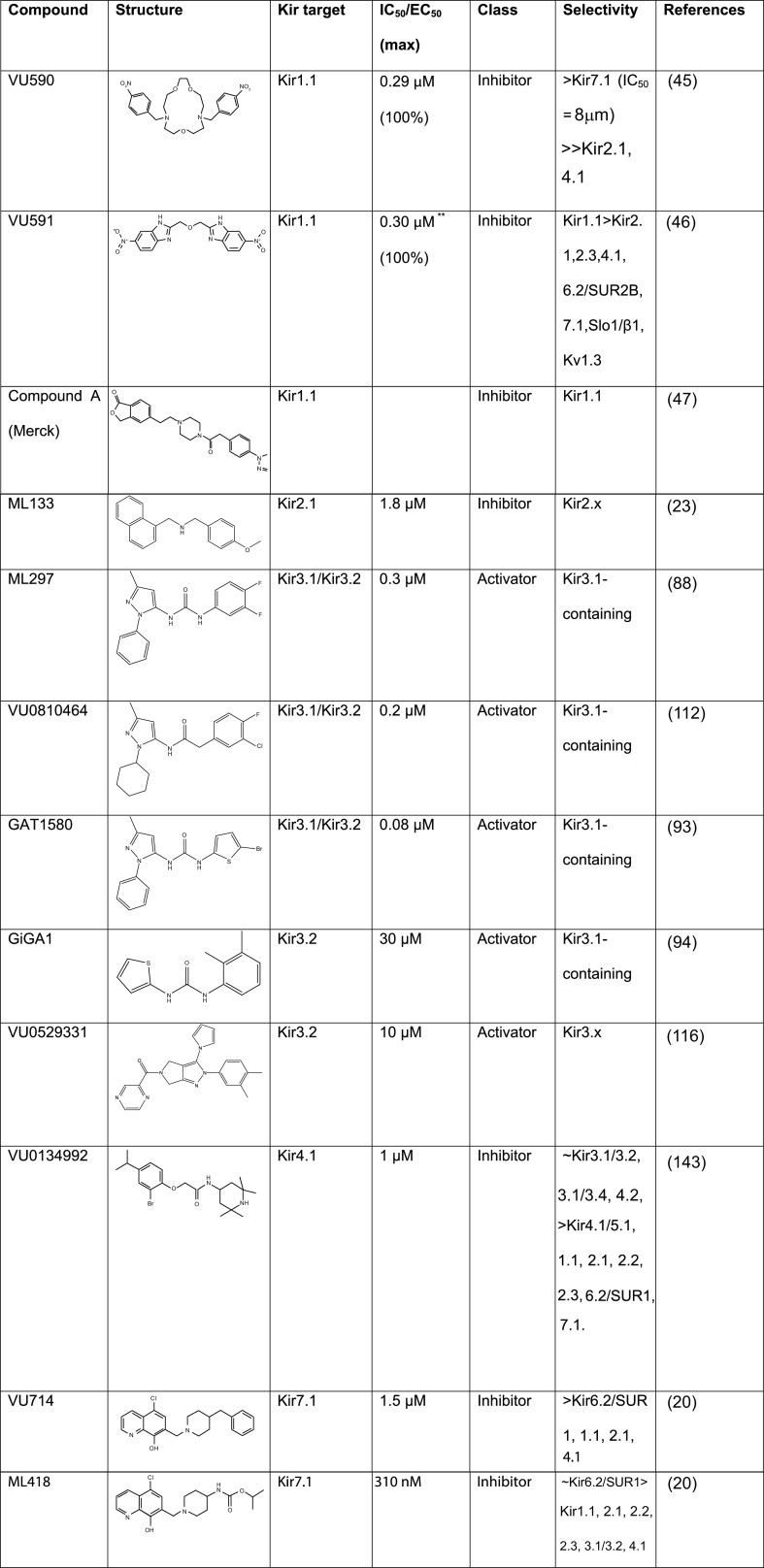
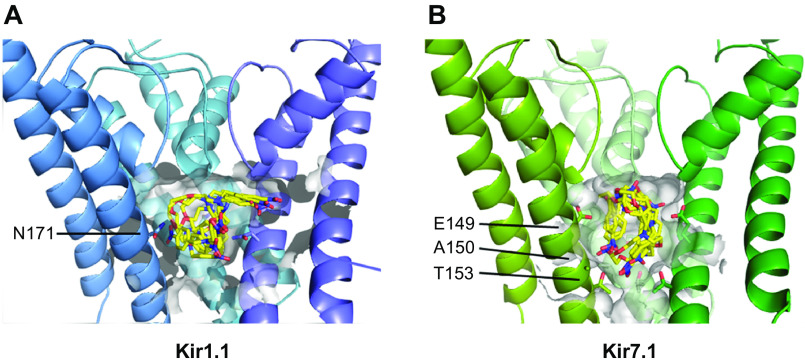
Inward rectifying potassium (Kir) channels play important roles in both excitable and nonexcitable cells of various organ systems and could represent valuable new drug targets for cardiovascular, metabolic, immune, and neurological diseases. In nonexcitable epithelial cells of the kidney tubule, for example, Kir1.1 (KCNJ1) and Kir4.1 (KCNJ10) are linked to sodium reabsorption in the thick ascending limb of Henle's loop and distal convoluted tubule, respectively, and have been explored as novel-mechanism diuretic targets for managing hypertension and edema. G protein-coupled Kir channels (Kir3) channels expressed in the central nervous system are critical effectors of numerous signal transduction pathways underlying analgesia, addiction, and respiratory-depressive effects of opioids. The historical dearth of pharmacological tool compounds for exploring the therapeutic potential of Kir channels has led to a molecular target-based approach using high-throughput screen (HTS) of small-molecule libraries and medicinal chemistry to develop "next-generation" Kir channel modulators that are both potent and specific for their targets. In this article, we review recent efforts focused specifically on discovery and improvement of target-selective molecular probes. The reader is introduced to fluorescence-based thallium flux assays that have enabled much of this work and then provided with an overview of progress made toward developing modulators of Kir1.1 (VU590, VU591), Kir2.x (ML133), Kir3.X (ML297, GAT1508, GiGA1, VU059331), Kir4.1 (VU0134992), and Kir7.1 (ML418). We discuss what is known about the small molecules' molecular mechanisms of action, in vitro and in vivo pharmacology, and then close with our view of what critical work remains to be done.
Keywords: drug discovery; high-throughput screening; medicinal chemistry; small molecules.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
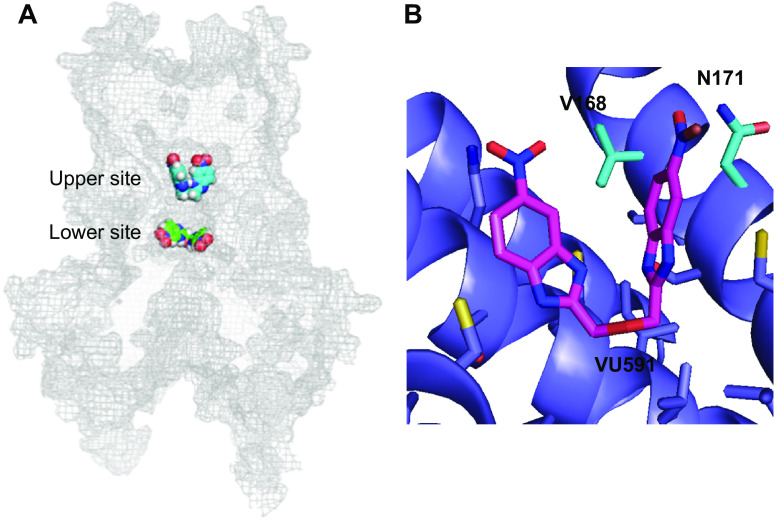
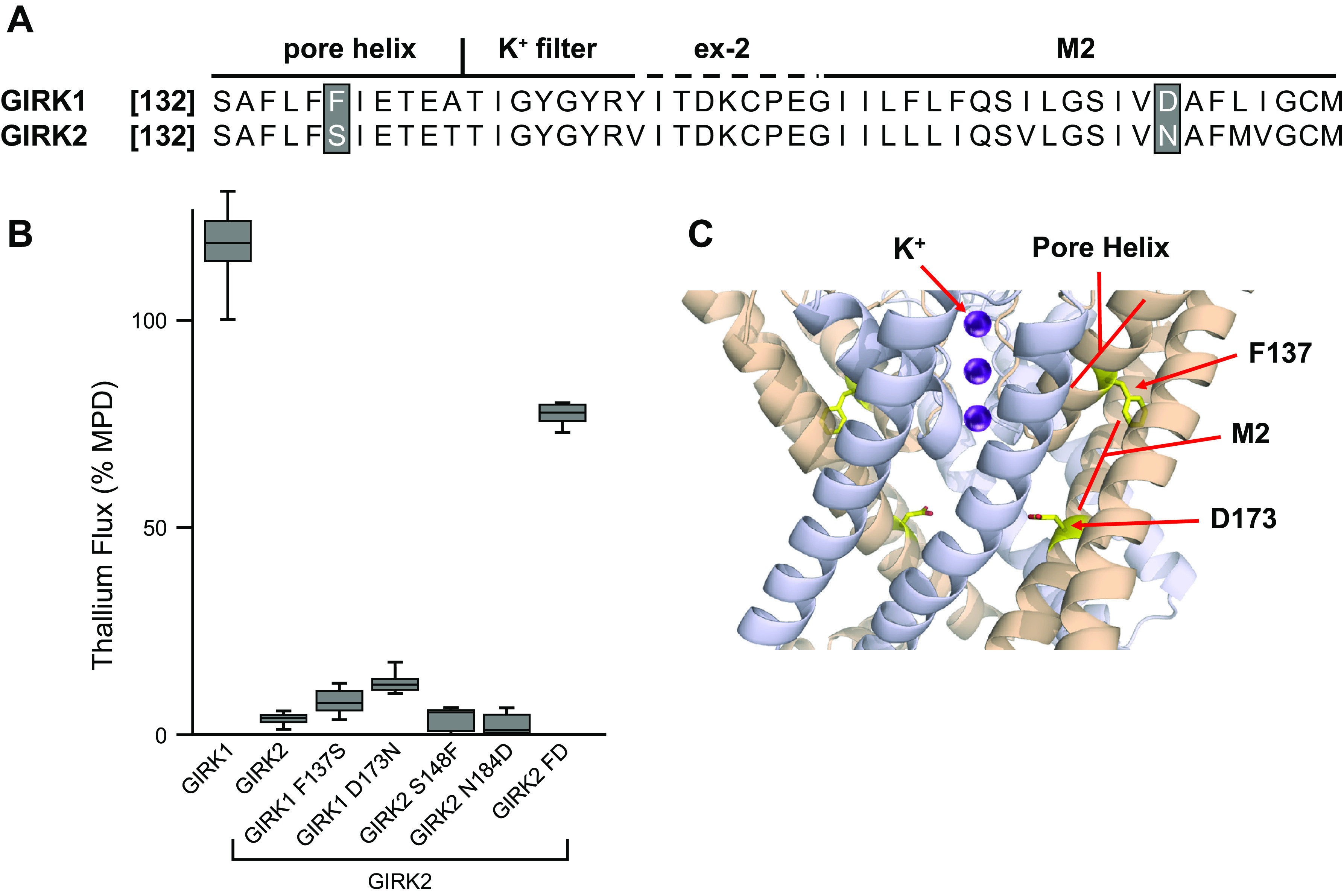
Similar articles
-
Discovery, Characterization, and Effects on Renal Fluid and Electrolyte Excretion of the Kir4.1 Potassium Channel Pore Blocker, VU0134992.Mol Pharmacol. 2018 Aug;94(2):926-937. doi: 10.1124/mol.118.112359. Epub 2018 Jun 12. Mol Pharmacol. 2018. PMID: 29895592 Free PMC article.
-
Screening Technologies for Inward Rectifier Potassium Channels: Discovery of New Blockers and Activators.SLAS Discov. 2020 Jun;25(5):420-433. doi: 10.1177/2472555220905558. Epub 2020 Apr 15. SLAS Discov. 2020. PMID: 32292089
-
High-throughput screening for small-molecule modulators of inward rectifier potassium channels.J Vis Exp. 2013 Jan 27;(71):4209. doi: 10.3791/4209. J Vis Exp. 2013. PMID: 23381507 Free PMC article.
-
Small-molecule modulators of inward rectifier K+ channels: recent advances and future possibilities.Future Med Chem. 2010 May;2(5):757-74. doi: 10.4155/fmc.10.179. Future Med Chem. 2010. PMID: 20543968 Free PMC article. Review.
-
Cardiac and renal inward rectifier potassium channel pharmacology: emerging tools for integrative physiology and therapeutics.Curr Opin Pharmacol. 2014 Apr;15:7-15. doi: 10.1016/j.coph.2013.11.002. Epub 2013 Nov 26. Curr Opin Pharmacol. 2014. PMID: 24721648 Free PMC article. Review.
Cited by
-
Kir4.1 is specifically expressed and active in non-myelinating Schwann cells.Glia. 2023 Apr;71(4):926-944. doi: 10.1002/glia.24315. Epub 2022 Dec 8. Glia. 2023. PMID: 36479906 Free PMC article.
-
Kir5.1 channels: potential role in epilepsy and seizure disorders.Am J Physiol Cell Physiol. 2022 Sep 1;323(3):C706-C717. doi: 10.1152/ajpcell.00235.2022. Epub 2022 Jul 18. Am J Physiol Cell Physiol. 2022. PMID: 35848616 Free PMC article. Review.
-
Consequences of somatic mutations of GIRK1 detected in primary malign tumors on expression and function of G-protein activated, inwardly rectifying, K+ channels.Front Oncol. 2022 Oct 31;12:998907. doi: 10.3389/fonc.2022.998907. eCollection 2022. Front Oncol. 2022. PMID: 36483038 Free PMC article.
-
Alternative Targets for Modulators of Mitochondrial Potassium Channels.Molecules. 2022 Jan 4;27(1):299. doi: 10.3390/molecules27010299. Molecules. 2022. PMID: 35011530 Free PMC article. Review.
-
K+ -independent Kir blockade by external Cs+ and Ba2.Physiol Rep. 2022 Mar;10(5):e15200. doi: 10.14814/phy2.15200. Physiol Rep. 2022. PMID: 35274814 Free PMC article.
References
-
- Nichols CG, Kj Enkvetchakul D, Flagg TP. KATP channels: from structure to disease. Biological Membranes 23: 101–110, 2006. https://www.researchgate.net/publication/283146317_KATP_channels_From_st....
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical