

Chaperoning SNARE Folding and Assembly
- PMID: 33823650
- PMCID: PMC8900292
- DOI: 10.1146/annurev-biochem-081820-103615
Chaperoning SNARE Folding and Assembly
Abstract
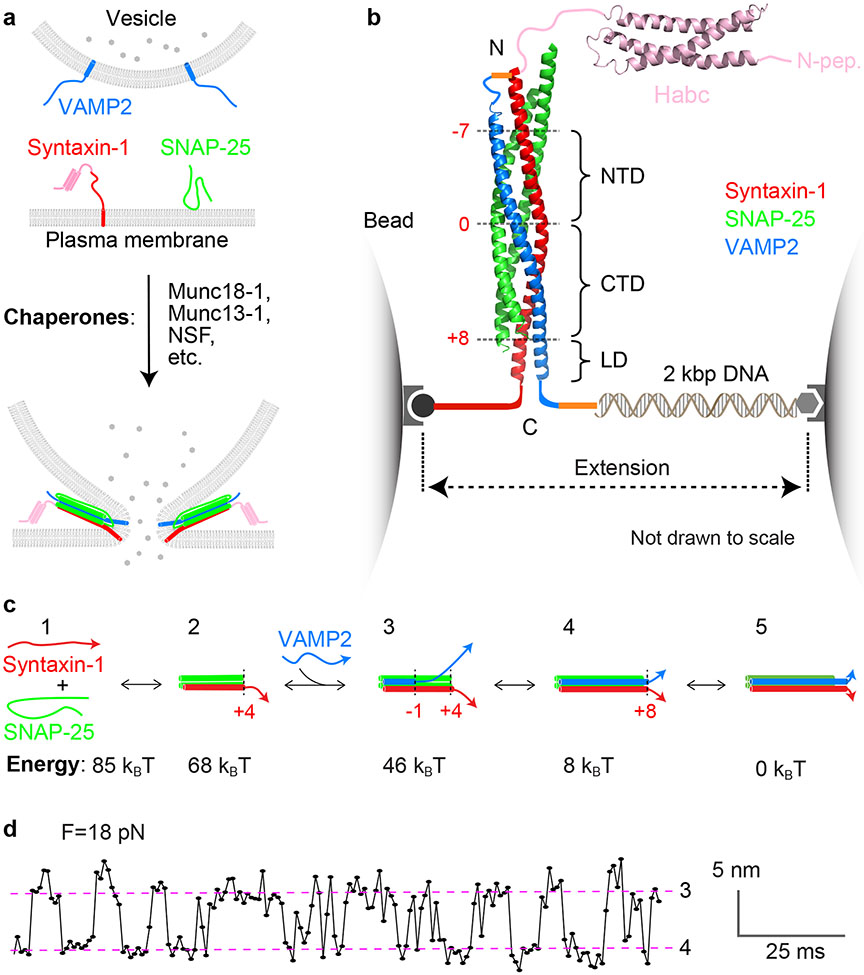
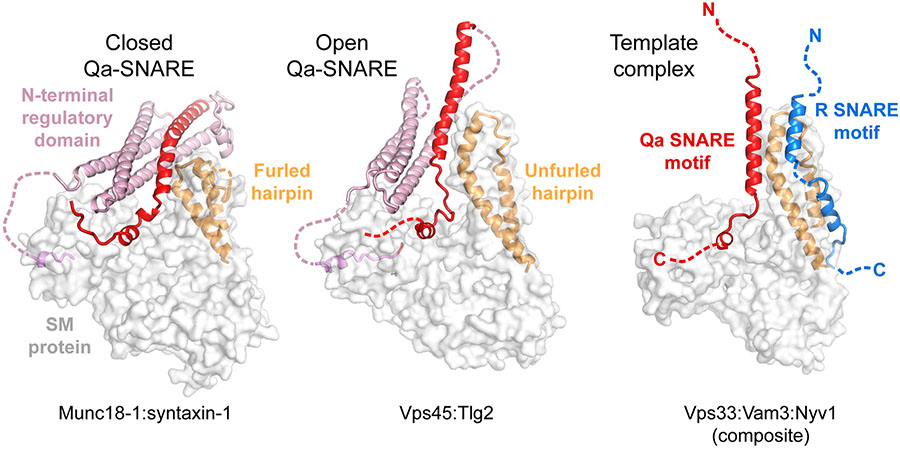
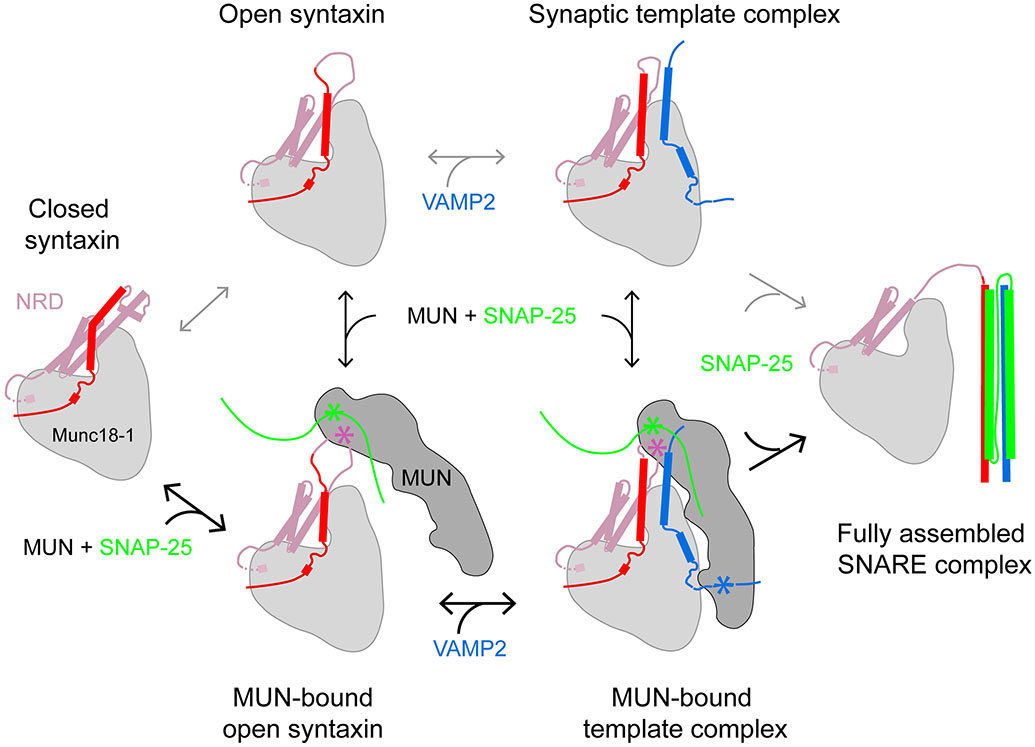
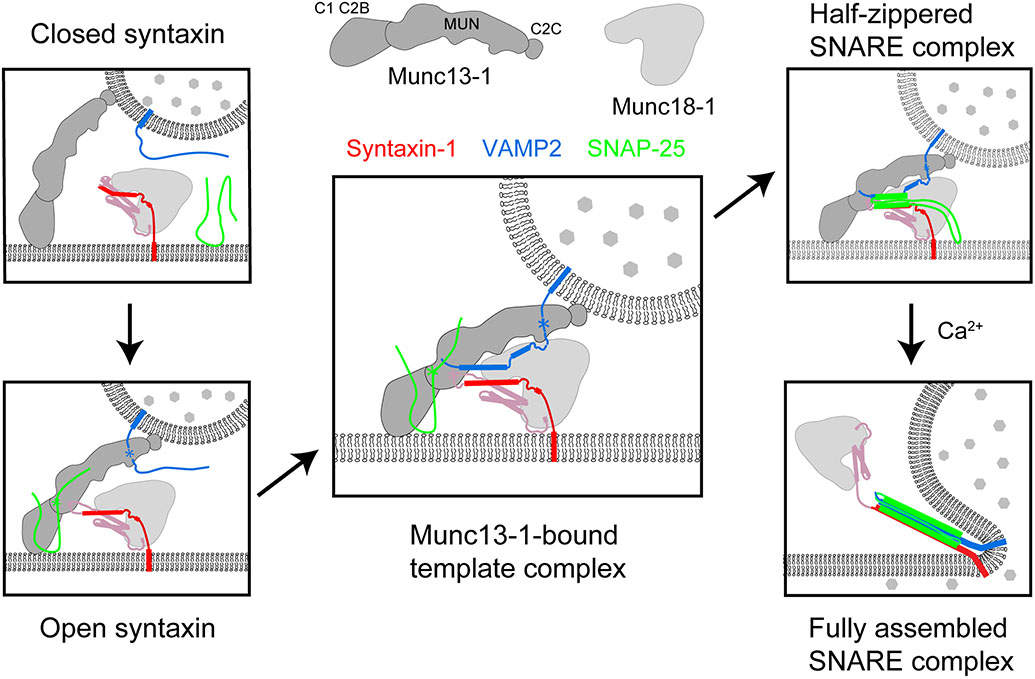
SNARE proteins and Sec1/Munc18 (SM) proteins constitute the core molecular engine that drives nearly all intracellular membrane fusion and exocytosis. While SNAREs are known to couple their folding and assembly to membrane fusion, the physiological pathways of SNARE assembly and the mechanistic roles of SM proteins have long been enigmatic. Here, we review recent advances in understanding the SNARE-SM fusion machinery with an emphasis on biochemical and biophysical studies of proteins that mediate synaptic vesicle fusion. We begin by discussing the energetics, pathways, and kinetics of SNARE folding and assembly in vitro. Then, we describe diverse interactions between SM and SNARE proteins and their potential impact on SNARE assembly in vivo. Recent work provides strong support for the idea that SM proteins function as chaperones, their essential role being to enable fast, accurate SNARE assembly. Finally, we review the evidence that SM proteins collaborate with other SNARE chaperones, especially Munc13-1, and briefly discuss some roles of SNARE and SM protein deficiencies in human disease.
Keywords: Munc18-1; SM proteins; SNARE assembly; membrane fusion; optical tweezers; template complex.
Figures
Similar articles
-
Munc13-1 MUN domain and Munc18-1 cooperatively chaperone SNARE assembly through a tetrameric complex.Proc Natl Acad Sci U S A. 2020 Jan 14;117(2):1036-1041. doi: 10.1073/pnas.1914361117. Epub 2019 Dec 30. Proc Natl Acad Sci U S A. 2020. PMID: 31888993 Free PMC article.
-
Single-Molecule Manipulation Study of Chaperoned SNARE Folding and Assembly with Optical Tweezers.Methods Mol Biol. 2022;2478:461-481. doi: 10.1007/978-1-0716-2229-2_17. Methods Mol Biol. 2022. PMID: 36063331
-
Syntaxin N-terminal peptide motif is an initiation factor for the assembly of the SNARE-Sec1/Munc18 membrane fusion complex.Proc Natl Acad Sci U S A. 2010 Dec 28;107(52):22399-406. doi: 10.1073/pnas.1012997108. Epub 2010 Dec 7. Proc Natl Acad Sci U S A. 2010. PMID: 21139055 Free PMC article.
-
Energetics, kinetics, and pathways of SNARE assembly in membrane fusion.Crit Rev Biochem Mol Biol. 2022 Aug;57(4):443-460. doi: 10.1080/10409238.2022.2121804. Epub 2022 Sep 24. Crit Rev Biochem Mol Biol. 2022. PMID: 36151854 Free PMC article. Review.
-
Energetics, kinetics, and pathway of SNARE folding and assembly revealed by optical tweezers.Protein Sci. 2017 Jul;26(7):1252-1265. doi: 10.1002/pro.3116. Epub 2017 Mar 8. Protein Sci. 2017. PMID: 28097727 Free PMC article. Review.
Cited by
-
The function of VAMP2 in mediating membrane fusion: An overview.Front Mol Neurosci. 2022 Dec 23;15:948160. doi: 10.3389/fnmol.2022.948160. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36618823 Free PMC article. Review.
-
Neuronal SNARE complex assembly guided by Munc18-1 and Munc13-1.FEBS Open Bio. 2022 Nov;12(11):1939-1957. doi: 10.1002/2211-5463.13394. Epub 2022 Mar 22. FEBS Open Bio. 2022. PMID: 35278279 Free PMC article. Review.
-
Mechanisms of SNARE proteins in membrane fusion.Nat Rev Mol Cell Biol. 2024 Feb;25(2):101-118. doi: 10.1038/s41580-023-00668-x. Epub 2023 Oct 17. Nat Rev Mol Cell Biol. 2024. PMID: 37848589 Free PMC article. Review.
-
Vesicle trafficking and vesicle fusion: mechanisms, biological functions, and their implications for potential disease therapy.Mol Biomed. 2022 Sep 21;3(1):29. doi: 10.1186/s43556-022-00090-3. Mol Biomed. 2022. PMID: 36129576 Free PMC article. Review.
-
On the difficulties of characterizing weak protein interactions that are critical for neurotransmitter release.FEBS Open Bio. 2022 Nov;12(11):1912-1938. doi: 10.1002/2211-5463.13473. Epub 2022 Sep 2. FEBS Open Bio. 2022. PMID: 35986639 Free PMC article. Review.
References
-
- Rothman JE. 2014. The principle of membrane fusion in the cell (Nobel lecture). Angew. Chem. Int. Ed 53:12676–94 - PubMed
-
- Südhof TC. 2014. The molecular machinery of neurotransmitter release (Nobel lecture). Angew. Chem. Int. Ed 53:12696–717 - PubMed
-
- Stamberger H, Nikanorova M, Willemsen MH, Accorsi P, Angriman M, et al. 2016. STXBP1 encephalopathy: a neurodevelopmental disorder including epilepsy. Neurology 86:954–62 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
