

Chimeric Zika viruses containing structural protein genes of insect-specific flaviviruses cannot replicate in vertebrate cells due to entry and post-translational restrictions
- PMID: 33812340
- PMCID: PMC8205982
- DOI: 10.1016/j.virol.2021.03.014
Chimeric Zika viruses containing structural protein genes of insect-specific flaviviruses cannot replicate in vertebrate cells due to entry and post-translational restrictions
Abstract
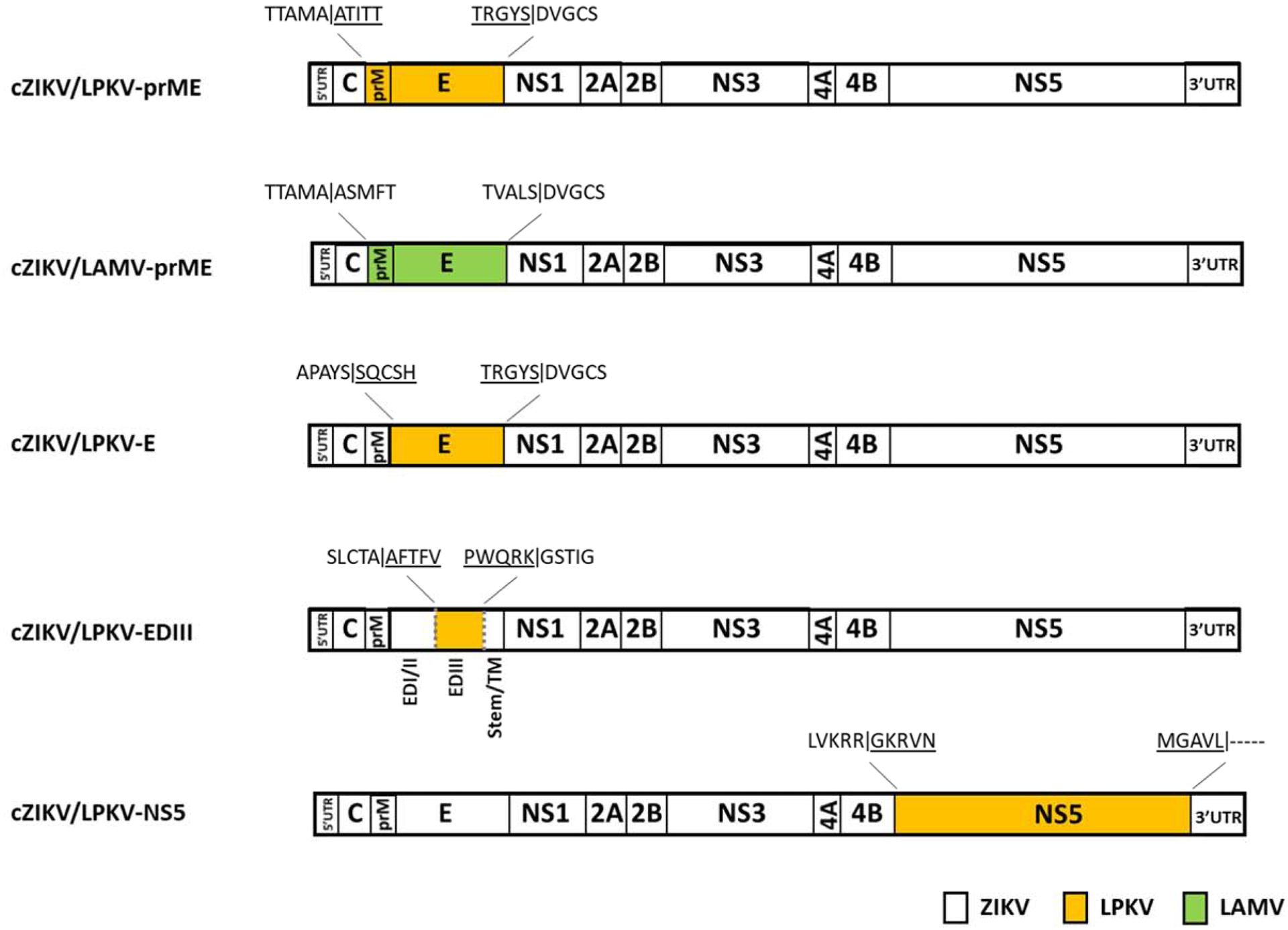
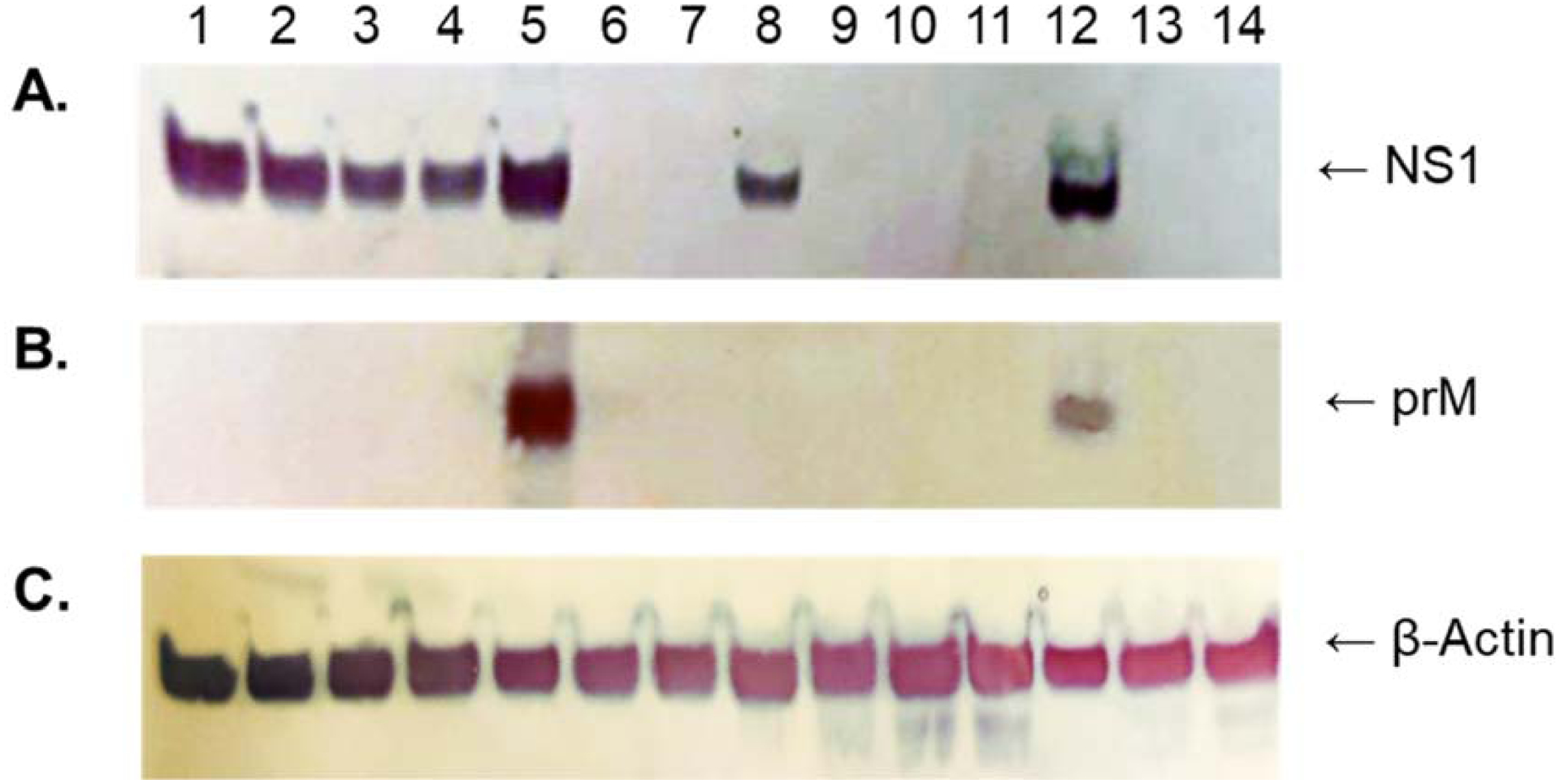
Long Pine Key virus (LPKV) and Lammi virus are insect-specific flaviviruses that phylogenetically affiliate with dual-host flaviviruses. The goal of this study was to provide insight into the genetic determinants that condition this host range restriction. Chimeras were initially created by replacing select regions of the Zika virus genome, including the premembrane and envelope protein (prM-E) genes, with the corresponding regions of the LPKV genome. Of the four chimeras produced, one (the prM-E swap) yielded virus that replicated in mosquito cells. Another chimeric virus with a mosquito replication-competent phenotype was created by inserting the prM-E genes of Lammi virus into a Zika virus genetic background. Vertebrate cells did not support the replication of either chimeric virus although trace to modest amounts of viral antigen were produced, consistent with suboptimal viral entry. These data suggest that dual-host affiliated insect-specific flaviviruses cannot replicate in vertebrate cells due to entry and post-translational restrictions.
Keywords: Chimeric; Entry; Flavivirus; Host range; Insect-specific.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
COMPETING INTERESTS
None of declare
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

Similar articles
-
The host range restriction of bat-associated no-known-vector flaviviruses occurs post-entry.J Gen Virol. 2021 Sep;102(9):001647. doi: 10.1099/jgv.0.001647. J Gen Virol. 2021. PMID: 34486974 Free PMC article.
-
Evidence that untranslated genomic sequences are key determinants of insect-specific flavivirus host restriction.Virology. 2022 Sep;574:102-114. doi: 10.1016/j.virol.2022.07.013. Epub 2022 Aug 5. Virology. 2022. PMID: 35969917
-
Antigenic Characterization of New Lineage II Insect-Specific Flaviviruses in Australian Mosquitoes and Identification of Host Restriction Factors.mSphere. 2020 Jun 17;5(3):e00095-20. doi: 10.1128/mSphere.00095-20. mSphere. 2020. PMID: 32554715 Free PMC article.
-
Insect-specific flaviviruses: a systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization.Viruses. 2015 Apr 10;7(4):1927-59. doi: 10.3390/v7041927. Viruses. 2015. PMID: 25866904 Free PMC article. Review.
-
Establishment and Application of Flavivirus Replicons.Adv Exp Med Biol. 2018;1062:165-173. doi: 10.1007/978-981-10-8727-1_12. Adv Exp Med Biol. 2018. PMID: 29845532 Review.
Cited by
-
The host range restriction of bat-associated no-known-vector flaviviruses occurs post-entry.J Gen Virol. 2021 Sep;102(9):001647. doi: 10.1099/jgv.0.001647. J Gen Virol. 2021. PMID: 34486974 Free PMC article.
-
Replication is the key barrier during the dual-host adaptation of mosquito-borne flaviviruses.Proc Natl Acad Sci U S A. 2022 Mar 22;119(12):e2110491119. doi: 10.1073/pnas.2110491119. Epub 2022 Mar 16. Proc Natl Acad Sci U S A. 2022. PMID: 35294288 Free PMC article.
-
Isolation of a novel insect-specific flavivirus with immunomodulatory effects in vertebrate systems.Virology. 2021 Oct;562:50-62. doi: 10.1016/j.virol.2021.07.004. Epub 2021 Jul 8. Virology. 2021. PMID: 34256244 Free PMC article.
-
Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses.Vaccines (Basel). 2021 Oct 22;9(11):1230. doi: 10.3390/vaccines9111230. Vaccines (Basel). 2021. PMID: 34835160 Free PMC article. Review.
-
Production of a chimeric flavivirus that contains the major structural glycoprotein genes of T'Ho virus in the genetic background of Zika virus.Virol J. 2023 Sep 1;20(1):197. doi: 10.1186/s12985-023-02172-2. Virol J. 2023. PMID: 37658438 Free PMC article.
References
-
- Alsaleh K, Khou C, Frenkiel MP, Lecollinet S, Vazquez A, de Arellano ER, Despres P, Pardigon N, 2016. The E glycoprotein plays an essential role in the high pathogenicity of European-Mediterranean IS98 strain of West Nile virus. Virology 492, 53–65. - PubMed
-
- Bhuvanakantham R, Cheong YK, Ng ML, 2010. West Nile virus capsid protein interaction with importin and HDM2 protein is regulated by protein kinase C-mediated phosphorylation. Microbes Infect 12, 615–625. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
