

Involvement of ethylene receptors in the salt tolerance response of Cucurbita pepo
- PMID: 33790231
- PMCID: PMC8012379
- DOI: 10.1038/s41438-021-00508-z
Involvement of ethylene receptors in the salt tolerance response of Cucurbita pepo
Abstract
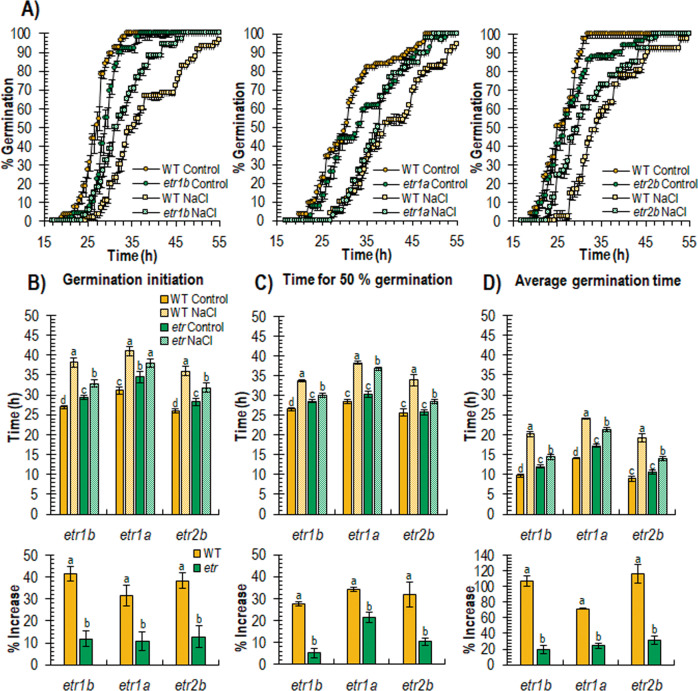
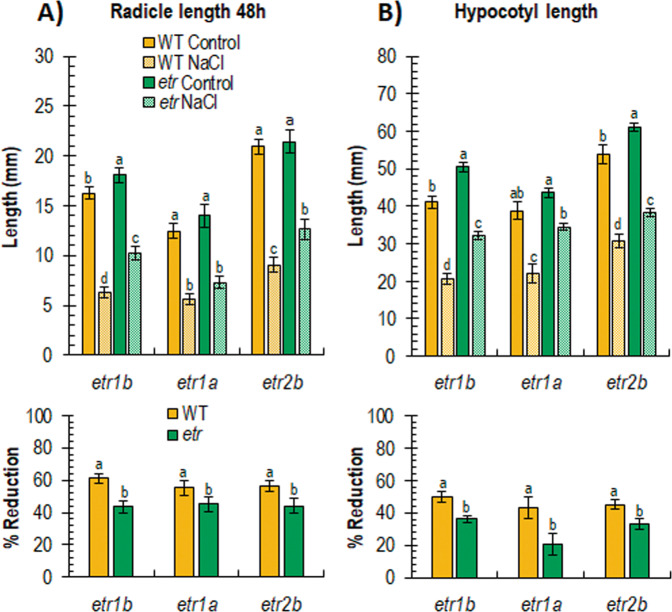
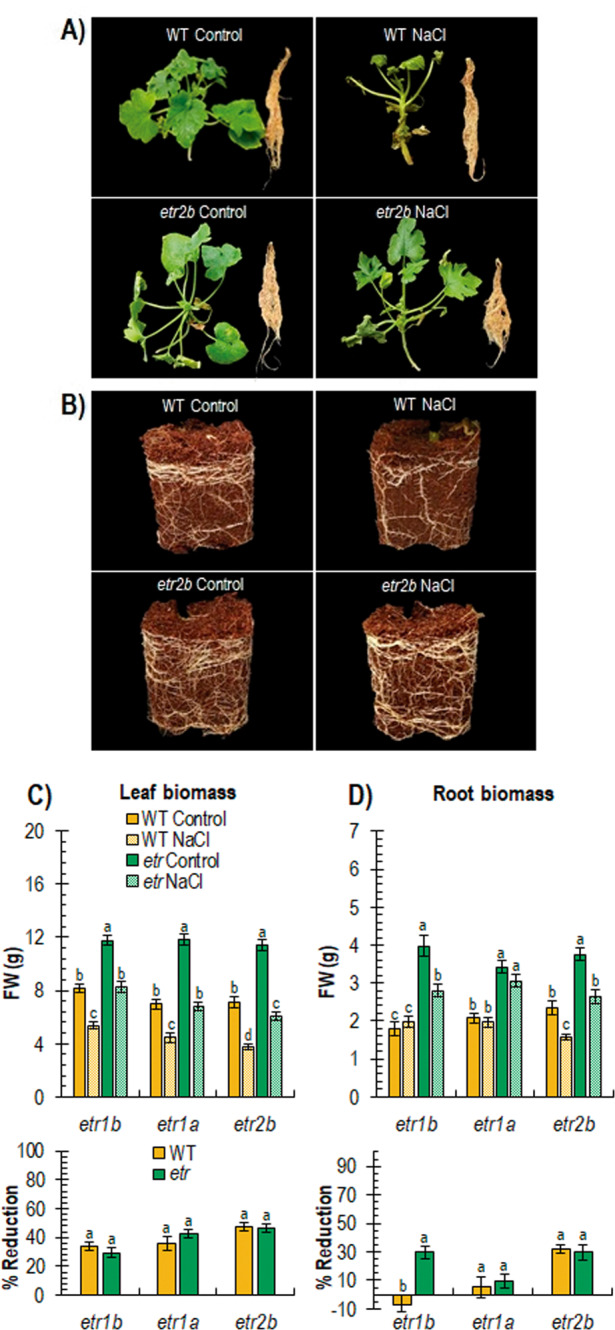
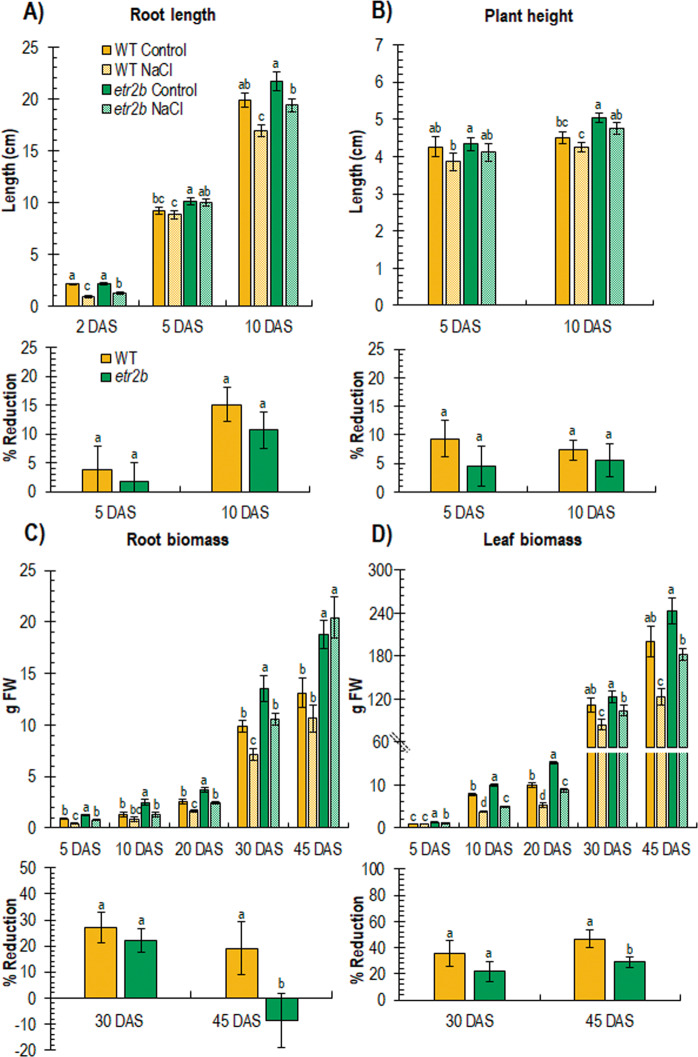
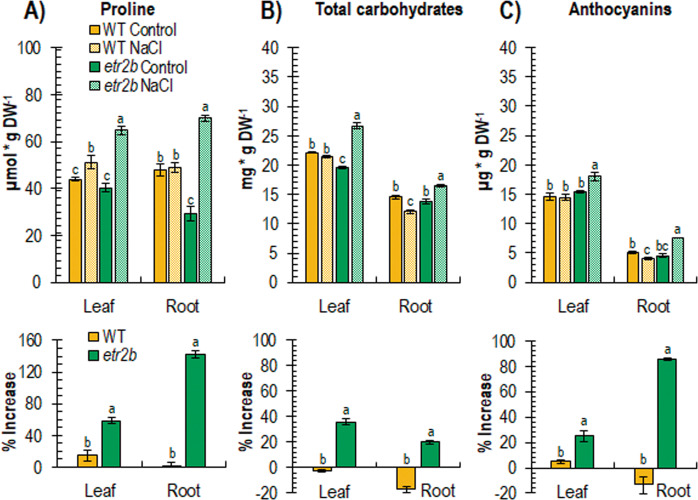
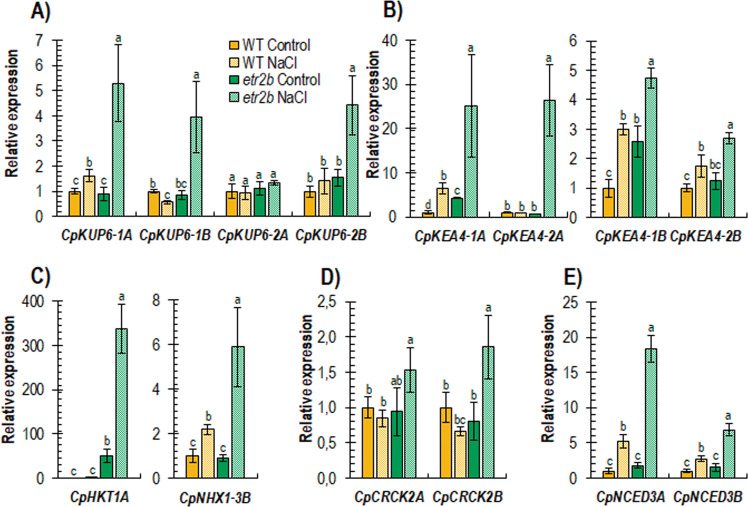
Abiotic stresses have a negative effect on crop production, affecting both vegetative and reproductive development. Ethylene plays a relevant role in plant response to environmental stresses, but the specific contribution of ethylene biosynthesis and signalling components in the salt stress response differs between Arabidopsis and rice, the two most studied model plants. In this paper, we study the effect of three gain-of-function mutations affecting the ethylene receptors CpETR1B, CpETR1A, and CpETR2B of Cucurbita pepo on salt stress response during germination, seedling establishment, and subsequent vegetative growth of plants. The mutations all reduced ethylene sensitivity, but enhanced salt tolerance, during both germination and vegetative growth, demonstrating that the three ethylene receptors play a positive role in salt tolerance. Under salt stress, etr1b, etr1a, and etr2b germinate earlier than WT, and the root and shoot growth rates of both seedlings and plants were less affected in mutant than in WT. The enhanced salt tolerance response of the etr2b plants was associated with a reduced accumulation of Na+ in shoots and leaves, as well as with a higher accumulation of compatible solutes, including proline and total carbohydrates, and antioxidant compounds, such as anthocyanin. Many membrane monovalent cation transporters, including Na+/H+ and K+/H+ exchangers (NHXs), K+ efflux antiporters (KEAs), high-affinity K+ transporters (HKTs), and K+ uptake transporters (KUPs) were also highly upregulated by salt in etr2b in comparison with WT. In aggregate, these data indicate that the enhanced salt tolerance of the mutant is led by the induction of genes that exclude Na+ in photosynthetic organs, while maintaining K+/Na+ homoeostasis and osmotic adjustment. If the salt response of etr mutants occurs via the ethylene signalling pathway, our data show that ethylene is a negative regulator of salt tolerance during germination and vegetative growth. Nevertheless, the higher upregulation of genes involved in Ca2+ signalling (CpCRCK2A and CpCRCK2B) and ABA biosynthesis (CpNCED3A and CpNCED3B) in etr2b leaves under salt stress likely indicates that the function of ethylene receptors in salt stress response in C. pepo can be mediated by Ca2+ and ABA signalling pathways.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Crosstalk between Ethylene, Jasmonate and ABA in Response to Salt Stress during Germination and Early Plant Growth in Cucurbita pepo.Int J Mol Sci. 2024 Aug 10;25(16):8728. doi: 10.3390/ijms25168728. Int J Mol Sci. 2024. PMID: 39201415 Free PMC article.
-
The ethylene receptor mutation etr2b reveals crosstalk between ethylene and ABA in the control of Cucurbita pepo germination.Physiol Plant. 2023 Jan;175(1):e13864. doi: 10.1111/ppl.13864. Physiol Plant. 2023. PMID: 36718078
-
Physiological and metabolomic responses of the ethylene insensitive squash mutant etr2b to drought.Plant Sci. 2023 Nov;336:111853. doi: 10.1016/j.plantsci.2023.111853. Epub 2023 Sep 1. Plant Sci. 2023. PMID: 37659732
-
The ethylene receptors CpETR1A and CpETR2B cooperate in the control of sex determination in Cucurbita pepo.J Exp Bot. 2020 Jan 1;71(1):154-167. doi: 10.1093/jxb/erz417. J Exp Bot. 2020. PMID: 31562498 Free PMC article.
-
Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge.Plants (Basel). 2022 Aug 25;11(17):2211. doi: 10.3390/plants11172211. Plants (Basel). 2022. PMID: 36079592 Free PMC article. Review.
Cited by
-
Crosstalk between Ethylene, Jasmonate and ABA in Response to Salt Stress during Germination and Early Plant Growth in Cucurbita pepo.Int J Mol Sci. 2024 Aug 10;25(16):8728. doi: 10.3390/ijms25168728. Int J Mol Sci. 2024. PMID: 39201415 Free PMC article.
-
A Comprehensive Evaluation of Salt Tolerance in Tomato (Var. Ailsa Craig): Responses of Physiological and Transcriptional Changes in RBOH's and ABA Biosynthesis and Signalling Genes.Int J Mol Sci. 2022 Jan 29;23(3):1603. doi: 10.3390/ijms23031603. Int J Mol Sci. 2022. PMID: 35163525 Free PMC article.
-
Research Advancements in Salt Tolerance of Cucurbitaceae: From Salt Response to Molecular Mechanisms.Int J Mol Sci. 2024 Aug 21;25(16):9051. doi: 10.3390/ijms25169051. Int J Mol Sci. 2024. PMID: 39201741 Free PMC article. Review.
-
A mutation in the brassinosteroid biosynthesis gene CpDWF5 disrupts vegetative and reproductive development and the salt stress response in squash (Cucurbita pepo).Hortic Res. 2024 Feb 23;11(4):uhae050. doi: 10.1093/hr/uhae050. eCollection 2024 Apr. Hortic Res. 2024. PMID: 38645681 Free PMC article.
-
Role of Ethylene Biosynthesis Genes in the Regulation of Salt Stress and Drought Stress Tolerance in Petunia.Front Plant Sci. 2022 Feb 23;13:844449. doi: 10.3389/fpls.2022.844449. eCollection 2022. Front Plant Sci. 2022. PMID: 35283920 Free PMC article.
References
-
- Zhang P, Zhang J, Chen M. Economic impacts of climate change on agriculture: the importance of additional climatic variables other than temperature and precipitation. J. Environ. Econ. Manag. 2016;83:8–31. doi: 10.1016/j.jeem.2016.12.001. - DOI
-
- Montanarella, L. et al (eds). Status of the World’s Soil Resources (Swsr)-main Report. (FAO, 2015).
-
- Gull, A., Ahmad Lone, A. & Ul Islam Wani, N. in Abiotic and Biotic Stress in Plants (ed. De Oliveira, A.) 3–9 (IntechOpen, 2019).
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
