

Conservation of the HBV RNA element epsilon in nackednaviruses reveals ancient origin of protein-primed reverse transcription
- PMID: 33753499
- PMCID: PMC8020639
- DOI: 10.1073/pnas.2022373118
Conservation of the HBV RNA element epsilon in nackednaviruses reveals ancient origin of protein-primed reverse transcription
Abstract
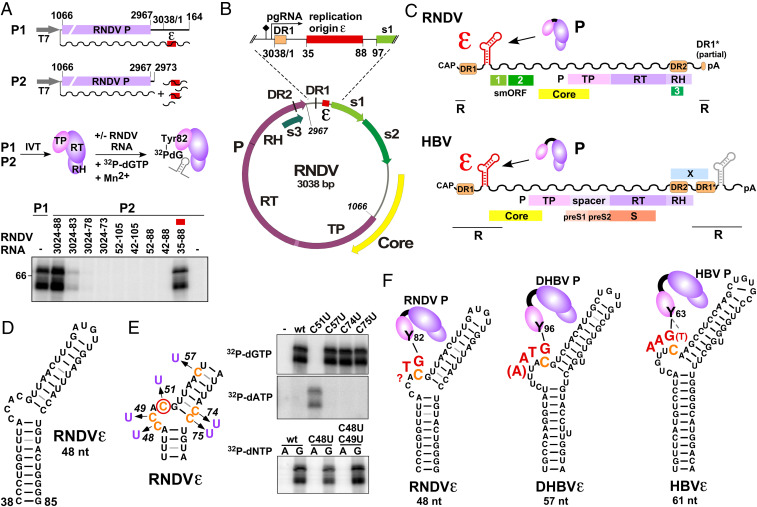
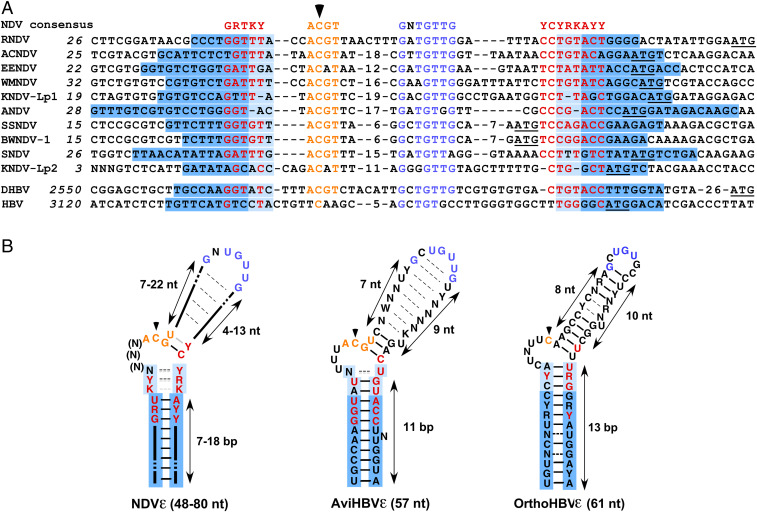
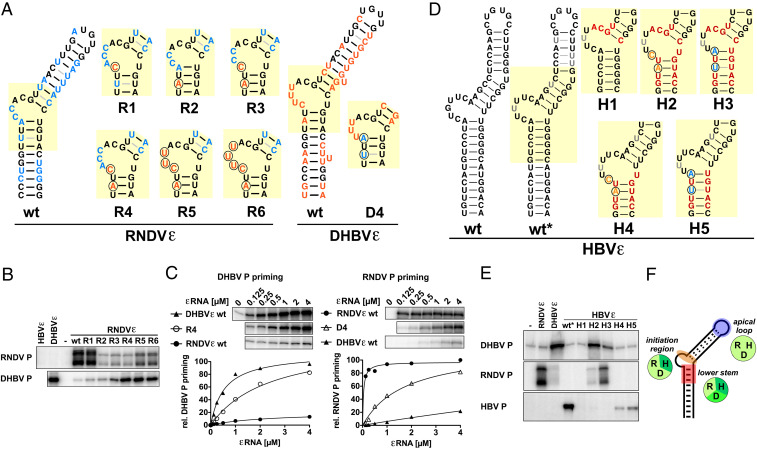
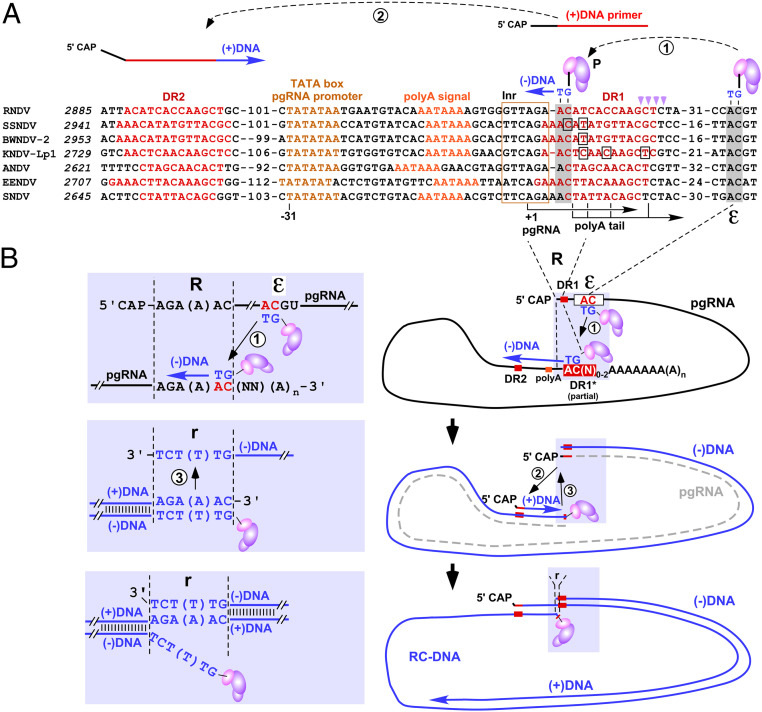
Hepadnaviruses, with the human hepatitis B virus as prototype, are small, enveloped hepatotropic DNA viruses which replicate by reverse transcription of an RNA intermediate. Replication is initiated by a unique protein-priming mechanism whereby a hydroxy amino acid side chain of the terminal protein (TP) domain of the viral polymerase (P) is extended into a short DNA oligonucleotide, which subsequently serves as primer for first-strand synthesis. A key component in the priming of reverse transcription is the viral RNA element epsilon, which contains the replication origin and serves as a template for DNA primer synthesis. Here, we show that recently discovered non-enveloped fish viruses, termed nackednaviruses [C. Lauber et al., Cell Host Microbe 22, 387-399 (2017)], employ a fundamentally similar replication mechanism despite their huge phylogenetic distance and major differences in genome organization and viral lifestyle. In vitro cross-priming studies revealed that few strategic nucleotide substitutions in epsilon enable site-specific protein priming by heterologous P proteins, demonstrating that epsilon is functionally conserved since the two virus families diverged more than 400 Mya. In addition, other cis elements crucial for the hepadnavirus-typical replication of pregenomic RNA into relaxed circular double-stranded DNA were identified at conserved positions in the nackednavirus genomes. Hence, the replication mode of both hepadnaviruses and nackednaviruses was already established in their Paleozoic common ancestor, making it a truly ancient and evolutionary robust principle of genome replication that is more widespread than previously thought.
Keywords: HBV long-term evolution; HBV replication mechanism; initiation of reverse transcription; paleovirology; protein priming.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
A bulged region of the hepatitis B virus RNA encapsidation signal contains the replication origin for discontinuous first-strand DNA synthesis.J Virol. 1996 May;70(5):2764-73. doi: 10.1128/JVI.70.5.2764-2773.1996. J Virol. 1996. PMID: 8627750 Free PMC article.
-
Deciphering the Origin and Evolution of Hepatitis B Viruses by Means of a Family of Non-enveloped Fish Viruses.Cell Host Microbe. 2017 Sep 13;22(3):387-399.e6. doi: 10.1016/j.chom.2017.07.019. Epub 2017 Aug 31. Cell Host Microbe. 2017. PMID: 28867387 Free PMC article.
-
Distinct families of cis-acting RNA replication elements epsilon from hepatitis B viruses.RNA Biol. 2012 Feb;9(2):130-6. doi: 10.4161/rna.18649. Epub 2012 Feb 1. RNA Biol. 2012. PMID: 22418844 Free PMC article.
-
Hepatitis B viruses: reverse transcription a different way.Virus Res. 2008 Jun;134(1-2):235-49. doi: 10.1016/j.virusres.2007.12.024. Epub 2008 Mar 12. Virus Res. 2008. PMID: 18339439 Review.
-
Structure and function of the encapsidation signal of hepadnaviridae.J Viral Hepat. 1998 Nov;5(6):357-67. doi: 10.1046/j.1365-2893.1998.00124.x. J Viral Hepat. 1998. PMID: 9857345 Review.
Cited by
-
Structural conservation of HBV-like capsid proteins over hundreds of millions of years despite the shift from non-enveloped to enveloped life-style.Nat Commun. 2023 Mar 22;14(1):1574. doi: 10.1038/s41467-023-37068-w. Nat Commun. 2023. PMID: 36949039 Free PMC article.
-
The Hepatitis B Virus Nucleocapsid-Dynamic Compartment for Infectious Virus Production and New Antiviral Target.Biomedicines. 2021 Oct 29;9(11):1577. doi: 10.3390/biomedicines9111577. Biomedicines. 2021. PMID: 34829806 Free PMC article. Review.
-
Predicted structure of the hepatitis B virus polymerase reveals an ancient conserved protein fold.Protein Sci. 2022 Oct;31(10):e4421. doi: 10.1002/pro.4421. Protein Sci. 2022. PMID: 36173165 Free PMC article.
-
The scientific basis of combination therapy for chronic hepatitis B functional cure.Nat Rev Gastroenterol Hepatol. 2023 Apr;20(4):238-253. doi: 10.1038/s41575-022-00724-5. Epub 2023 Jan 11. Nat Rev Gastroenterol Hepatol. 2023. PMID: 36631717 Review.
-
Opportunities and Challenges of Data-Driven Virus Discovery.Biomolecules. 2022 Aug 4;12(8):1073. doi: 10.3390/biom12081073. Biomolecules. 2022. PMID: 36008967 Free PMC article.
References
-
- Nassal M., Hepatitis B viruses: Reverse transcription a different way. Virus Res. 134, 235–249 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
