

The McdAB system positions α-carboxysomes in proteobacteria
- PMID: 33638215
- PMCID: PMC8359340
- DOI: 10.1111/mmi.14708
The McdAB system positions α-carboxysomes in proteobacteria
Abstract
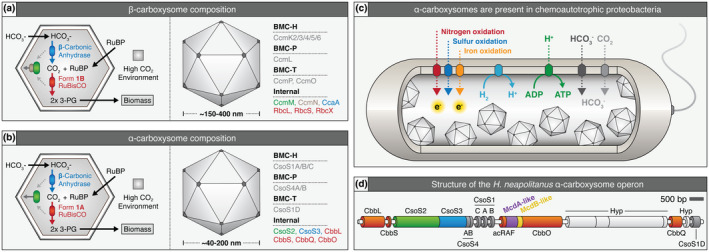
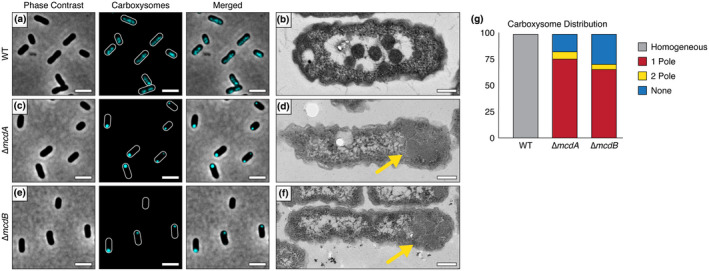
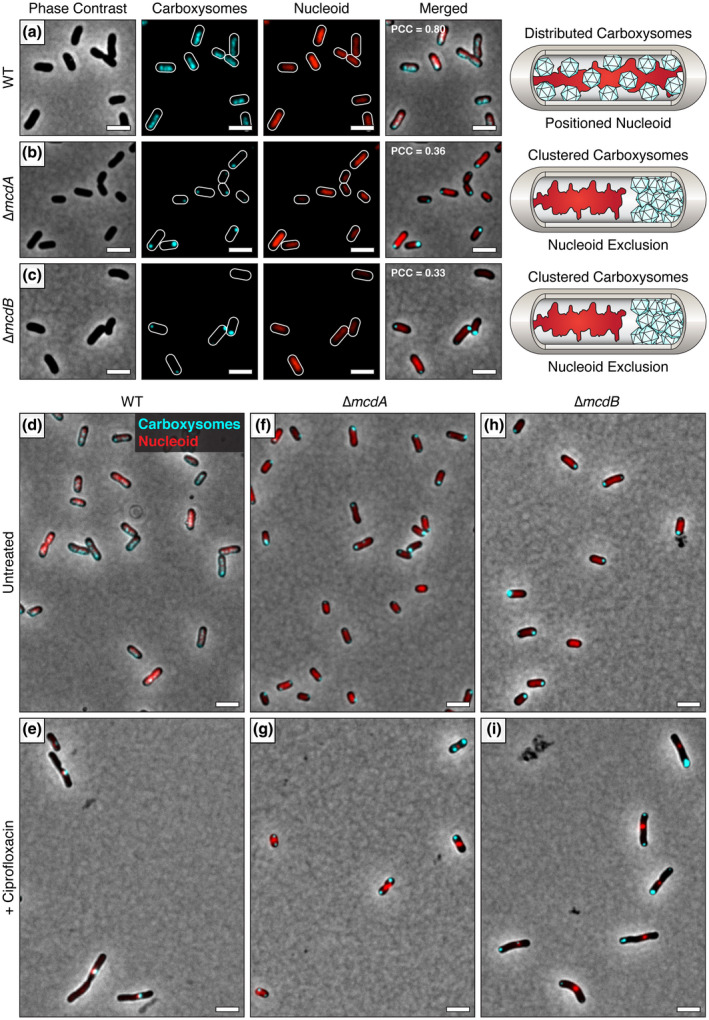
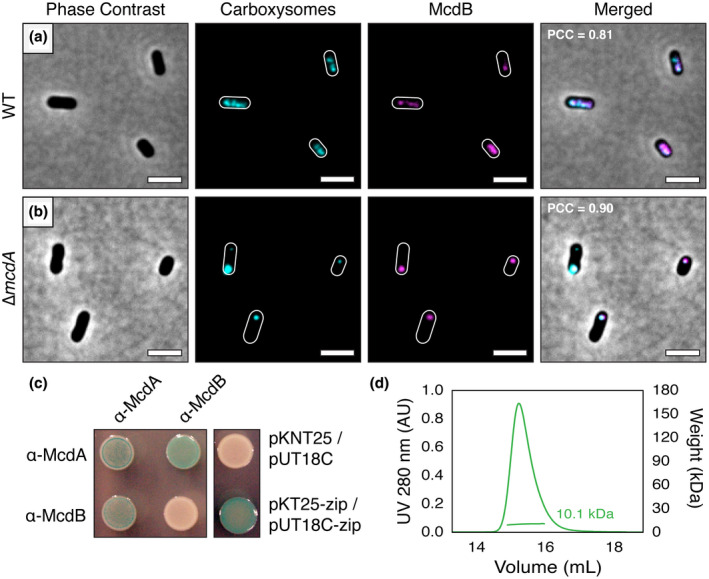
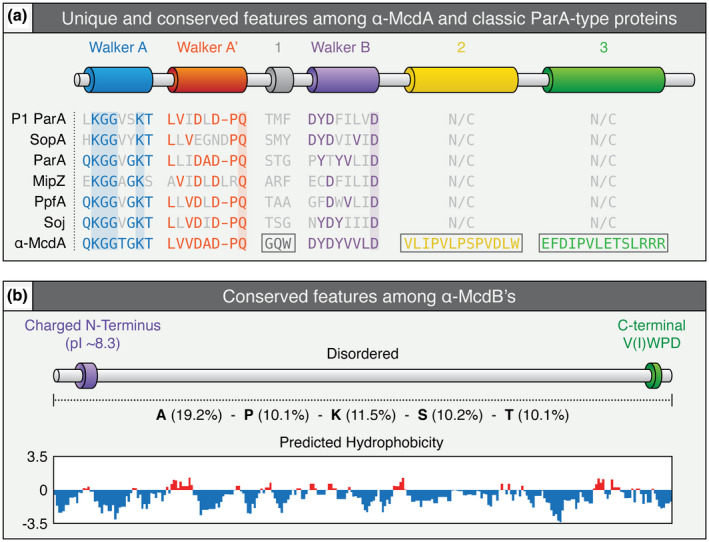
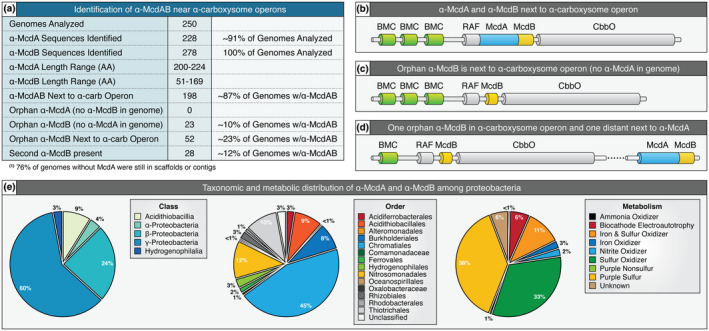
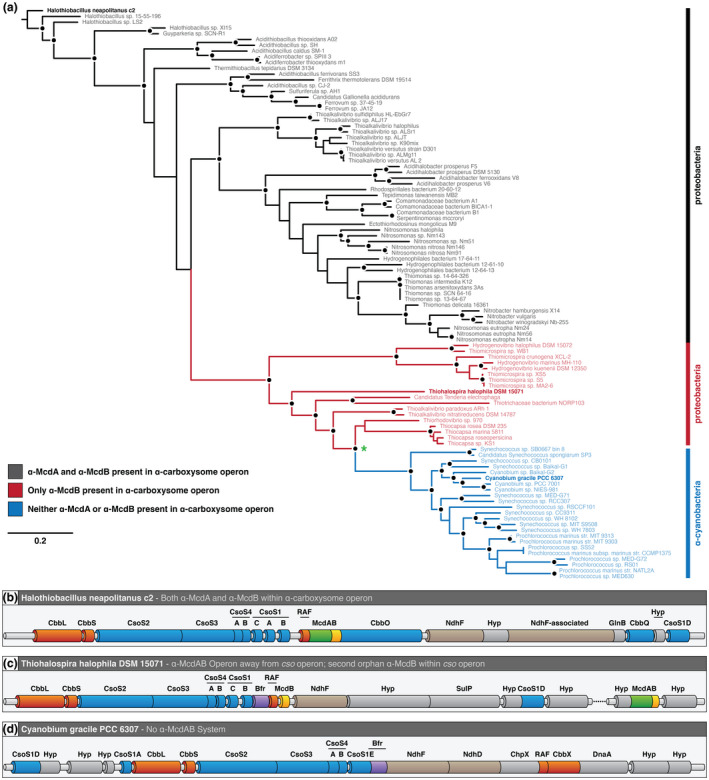
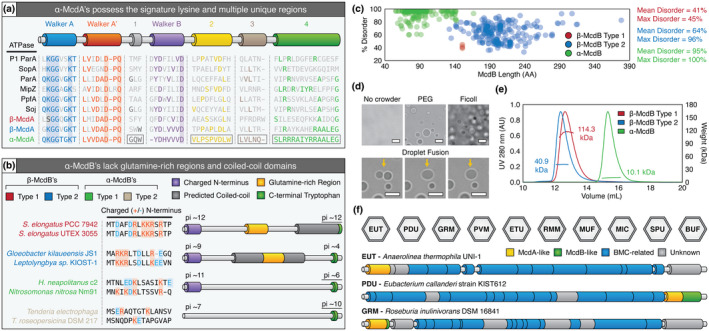
Carboxysomes are protein-based organelles essential for carbon fixation in cyanobacteria and proteobacteria. Previously, we showed that the cyanobacterial nucleoid is used to equally space out β-carboxysomes across cell lengths by a two-component system (McdAB) in the model cyanobacterium Synechococcus elongatus PCC 7942. More recently, we found that McdAB systems are widespread among β-cyanobacteria, which possess β-carboxysomes, but are absent in α-cyanobacteria, which possess structurally and phyletically distinct α-carboxysomes. Cyanobacterial α-carboxysomes are thought to have arisen in proteobacteria and then horizontally transferred into cyanobacteria, which suggests that α-carboxysomes in proteobacteria may also lack the McdAB system. Here, using the model chemoautotrophic proteobacterium Halothiobacillus neapolitanus, we show that a McdAB system distinct from that of β-cyanobacteria operates to position α-carboxysomes across cell lengths. We further show that this system is widespread among α-carboxysome-containing proteobacteria and that cyanobacteria likely inherited an α-carboxysome operon from a proteobacterium lacking the mcdAB locus. These results demonstrate that McdAB is a cross-phylum two-component system necessary for positioning both α- and β-carboxysomes. The findings have further implications for understanding the positioning of other protein-based bacterial organelles involved in diverse metabolic processes. PLAIN LANGUAGE SUMMARY: Cyanobacteria are well known to fix atmospheric CO2 into sugars using the enzyme Rubisco. Less appreciated are the carbon-fixing abilities of proteobacteria with diverse metabolisms. Bacterial Rubisco is housed within organelles called carboxysomes that increase enzymatic efficiency. Here we show that proteobacterial carboxysomes are distributed in the cell by two proteins, McdA and McdB. McdA on the nucleoid interacts with McdB on carboxysomes to equidistantly space carboxysomes from one another, ensuring metabolic homeostasis and a proper inheritance of carboxysomes following cell division. This study illuminates how widespread carboxysome positioning systems are among diverse bacteria. Carboxysomes significantly contribute to global carbon fixation; therefore, understanding the spatial organization mechanism shared across the bacterial world is of great interest.
© 2021 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Dissecting the phase separation and oligomerization activities of the carboxysome positioning protein McdB.Elife. 2023 Sep 5;12:e81362. doi: 10.7554/eLife.81362. Elife. 2023. PMID: 37668016 Free PMC article.
-
Origin and Evolution of Carboxysome Positioning Systems in Cyanobacteria.Mol Biol Evol. 2020 May 1;37(5):1434-1451. doi: 10.1093/molbev/msz308. Mol Biol Evol. 2020. PMID: 31899489 Free PMC article.
-
Carboxysome Mispositioning Alters Growth, Morphology, and Rubisco Level of the Cyanobacterium Synechococcus elongatus PCC 7942.mBio. 2021 Aug 31;12(4):e0269620. doi: 10.1128/mBio.02696-20. Epub 2021 Aug 3. mBio. 2021. PMID: 34340540 Free PMC article.
-
Cyanobacterial carboxysomes: microcompartments that facilitate CO2 fixation.J Mol Microbiol Biotechnol. 2013;23(4-5):300-7. doi: 10.1159/000351342. Epub 2013 Aug 5. J Mol Microbiol Biotechnol. 2013. PMID: 23920493 Review.
-
Advances in the bacterial organelles for CO2 fixation.Trends Microbiol. 2022 Jun;30(6):567-580. doi: 10.1016/j.tim.2021.10.004. Epub 2021 Nov 19. Trends Microbiol. 2022. PMID: 34802870 Review.
Cited by
-
Adapting from Low to High: An Update to CO2-Concentrating Mechanisms of Cyanobacteria and Microalgae.Plants (Basel). 2023 Apr 6;12(7):1569. doi: 10.3390/plants12071569. Plants (Basel). 2023. PMID: 37050194 Free PMC article. Review.
-
Positioning the Model Bacterial Organelle, the Carboxysome.mBio. 2021 May 11;12(3):e02519-19. doi: 10.1128/mBio.02519-19. mBio. 2021. PMID: 33975941 Free PMC article. Review.
-
The emergence of phase separation as an organizing principle in bacteria.Biophys J. 2021 Apr 6;120(7):1123-1138. doi: 10.1016/j.bpj.2020.09.023. Epub 2020 Sep 28. Biophys J. 2021. PMID: 33186556 Free PMC article. Review.
-
Dissecting the phase separation and oligomerization activities of the carboxysome positioning protein McdB.Elife. 2023 Sep 5;12:e81362. doi: 10.7554/eLife.81362. Elife. 2023. PMID: 37668016 Free PMC article.
-
Decoding the Absolute Stoichiometric Composition and Structural Plasticity of α-Carboxysomes.mBio. 2022 Apr 26;13(2):e0362921. doi: 10.1128/mbio.03629-21. Epub 2022 Mar 28. mBio. 2022. PMID: 35343789 Free PMC article.
References
-
- Alvarado, A., Kjæ, A., Yang, W., Mann, P., Briegel, A., Waldor, M.K. et al. (2017) Coupling chemosensory array formation and localization. eLife, 6. Available from: https://elifesciences.org/articles/31058 - PMC - PubMed
-
- Badger, M.R., Andrews, T.J., Whitney, S.M., Ludwig, M., Yellowlees, D.C., Leggat, W. et al. (1998) The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast‐based CO2‐concentrating mechanisms in algae. Canadian Journal of Botany, 76, 1052–1071. Available from: 10.1139/b98-074 - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
