

Vestigial mediates the effect of insulin signaling pathway on wing-morph switching in planthoppers
- PMID: 33561165
- PMCID: PMC7899339
- DOI: 10.1371/journal.pgen.1009312
Vestigial mediates the effect of insulin signaling pathway on wing-morph switching in planthoppers
Abstract
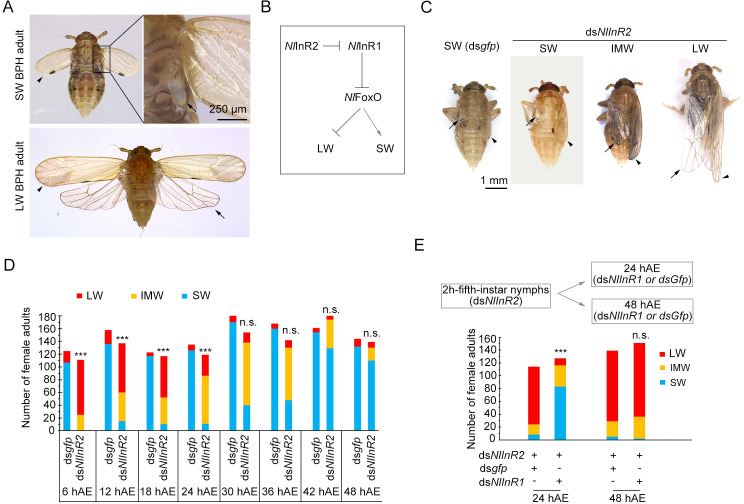
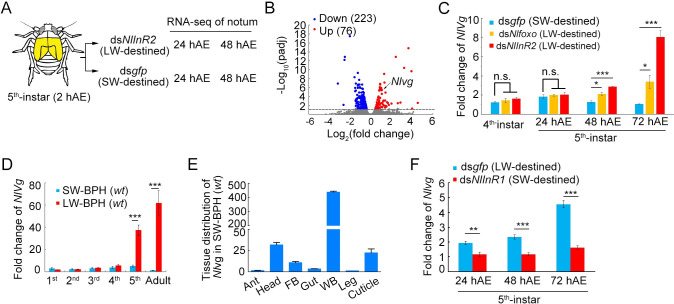
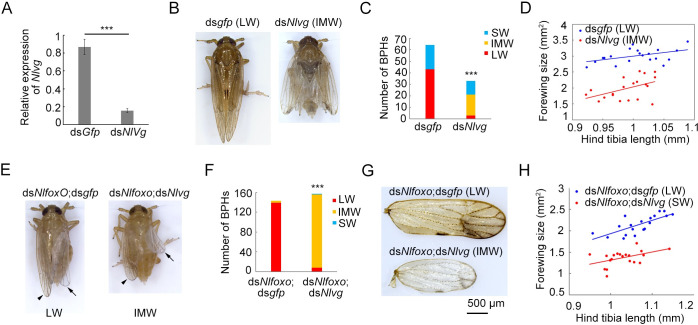
Wing polymorphism is an evolutionary feature found in a wide variety of insects, which offers a model system for studying the evolutionary significance of dispersal. In the wing-dimorphic planthopper Nilaparvata lugens, the insulin/insulin-like growth factor signaling (IIS) pathway acts as a 'master signal' that directs the development of either long-winged (LW) or short-winged (SW) morphs via regulation of the activity of Forkhead transcription factor subgroup O (NlFoxO). However, downstream effectors of the IIS-FoxO signaling cascade that mediate alternative wing morphs are unclear. Here we found that vestigial (Nlvg), a key wing-patterning gene, is selectively and temporally regulated by the IIS-FoxO signaling cascade during the wing-morph decision stage (fifth-instar stage). RNA interference (RNAi)-mediated silencing of Nlfoxo increase Nlvg expression in the fifth-instar stage (the last nymphal stage), thereby inducing LW development. Conversely, silencing of Nlvg can antagonize the effects of IIS activity on LW development, redirecting wing commitment from LW to the morph with intermediate wing size. In vitro and in vivo binding assays indicated that NlFoxO protein may suppress Nlvg expression by directly binding to the first intron region of the Nlvg locus. Our findings provide a first glimpse of the link connecting the IIS pathway to the wing-patterning network on the developmental plasticity of wings in insects, and help us understanding how phenotypic diversity is generated by the modification of a common set of pattern elements.
Conflict of interest statement
The authors declare that they have no competing interest.
Figures


Similar articles
-
Decapentaplegic function in wing vein development and wing morph transformation in brown planthopper, Nilaparvata lugens.Dev Biol. 2019 May 15;449(2):143-150. doi: 10.1016/j.ydbio.2019.02.016. Epub 2019 Mar 4. Dev Biol. 2019. PMID: 30844377
-
Two insulin receptors determine alternative wing morphs in planthoppers.Nature. 2015 Mar 26;519(7544):464-7. doi: 10.1038/nature14286. Epub 2015 Mar 18. Nature. 2015. PMID: 25799997
-
The transcription factor Zfh1 acts as a wing-morph switch in planthoppers.Nat Commun. 2022 Sep 27;13(1):5670. doi: 10.1038/s41467-022-33422-6. Nat Commun. 2022. PMID: 36167844 Free PMC article.
-
Insulin receptors and wing dimorphism in rice planthoppers.Philos Trans R Soc Lond B Biol Sci. 2017 Feb 5;372(1713):20150489. doi: 10.1098/rstb.2015.0489. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 27994130 Free PMC article. Review.
-
Molecular Mechanisms of Wing Polymorphism in Insects.Annu Rev Entomol. 2019 Jan 7;64:297-314. doi: 10.1146/annurev-ento-011118-112448. Epub 2018 Oct 12. Annu Rev Entomol. 2019. PMID: 30312555 Review.
Cited by
-
The diverse roles of insulin signaling in insect behavior.Front Insect Sci. 2024 Apr 4;4:1360320. doi: 10.3389/finsc.2024.1360320. eCollection 2024. Front Insect Sci. 2024. PMID: 38638680 Free PMC article. Review.
-
A plant virus manipulates the long-winged morph of insect vectors.Proc Natl Acad Sci U S A. 2024 Jan 16;121(3):e2315341121. doi: 10.1073/pnas.2315341121. Epub 2024 Jan 8. Proc Natl Acad Sci U S A. 2024. PMID: 38190519 Free PMC article.
-
A novel gene REPTOR2 activates the autophagic degradation of wing disc in pea aphid.Elife. 2023 Mar 21;12:e83023. doi: 10.7554/eLife.83023. Elife. 2023. PMID: 36943031 Free PMC article.
-
The vestigial Quadrant Enhancer is dispensable for pattern formation and development of the Drosophila wing.MicroPubl Biol. 2022 Jun 13;2022:10.17912/micropub.biology.000585. doi: 10.17912/micropub.biology.000585. eCollection 2022. MicroPubl Biol. 2022. PMID: 35783575 Free PMC article.
-
Neofunctionalization of a second insulin receptor gene in the wing-dimorphic planthopper, Nilaparvata lugens.PLoS Genet. 2021 Jun 28;17(6):e1009653. doi: 10.1371/journal.pgen.1009653. eCollection 2021 Jun. PLoS Genet. 2021. PMID: 34181658 Free PMC article.
References
-
- Harrison RG. Dispersal polymorphisms in insects. Annu Rev Ecol Syst. 1980;11:95–118. 10.1146/annurev.es.11.110180.000523 - DOI
-
- Roff DA. Life-history consequences of bioenergetic and biomechanical constraints on migration. Am Zool. 1991;31:205–15.
-
- Denno RF, Roderick GK. Density-related dispersal in planthoppers-Effects of interspecific crowding. Ecology. 1992;73:1323–34. 10.2307/1940679 - DOI
-
- Roff DA, Fairbairn DJ. Wing dimorphisms and the evolution of migratory polymorphisms among the insecta. Am Zool. 1991;31:243–51.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
